
Spawning Phenology and Early Growth of Japanese Anchovy (Engraulis japonicus) off the Pacific Coast of Japan
Abstract
:1. Introduction
2. Materials and Methods
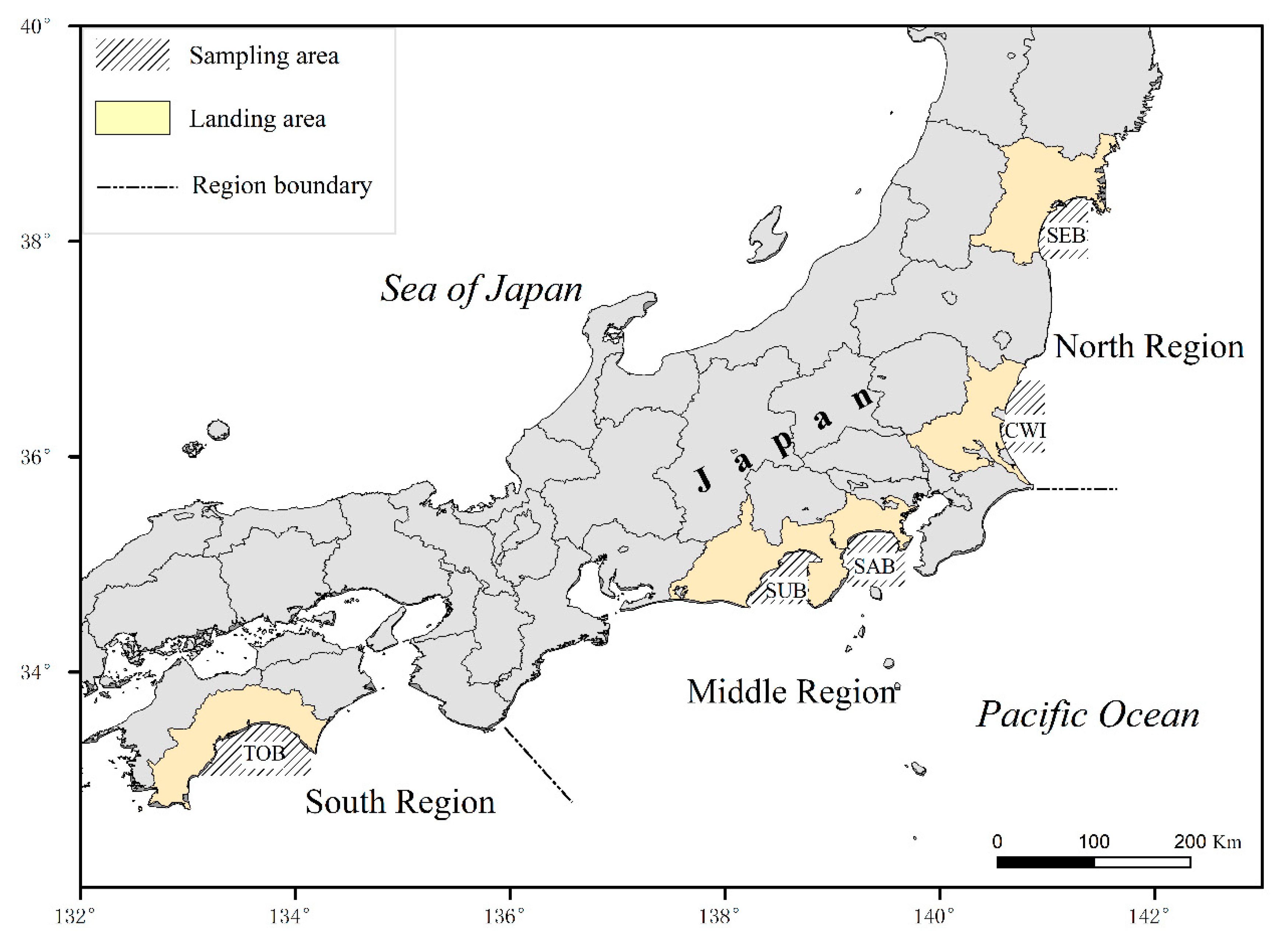
2.1. Sampling and Otolith Analysis
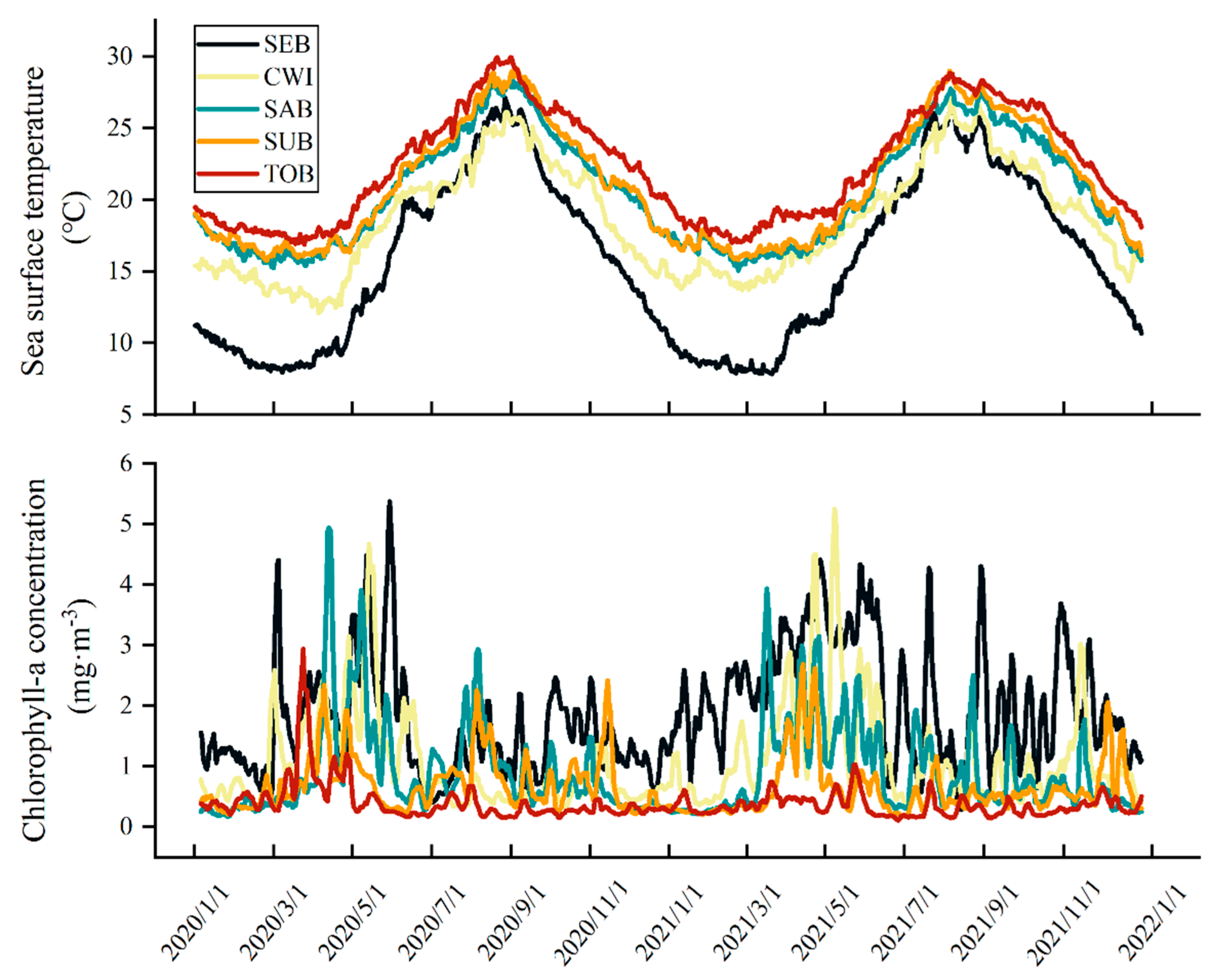
2.2. Aging and Environmental History Reconstruction
2.3. Growth Analysis and Modeling
3. Results
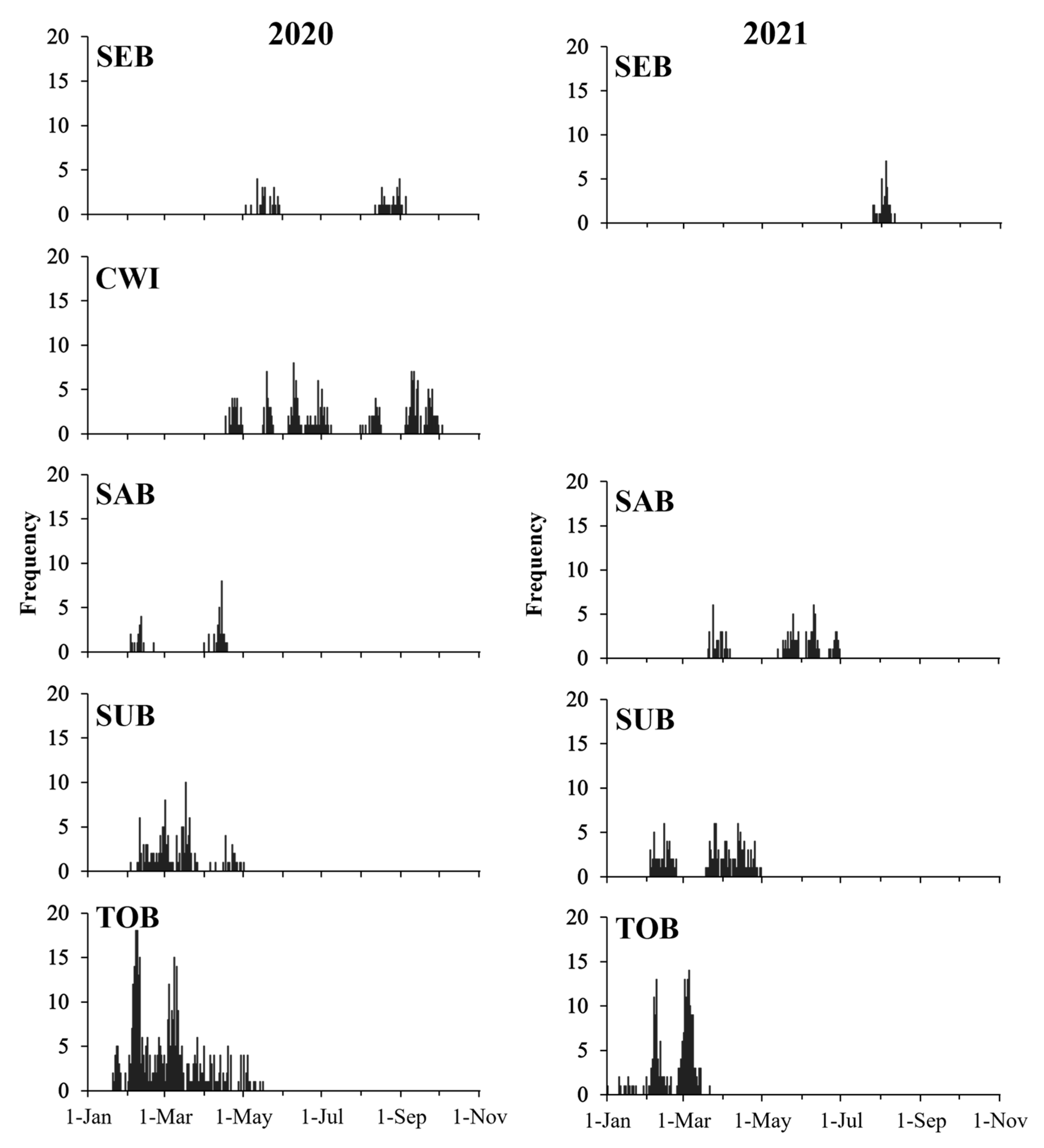
3.1. Spawning Phenology
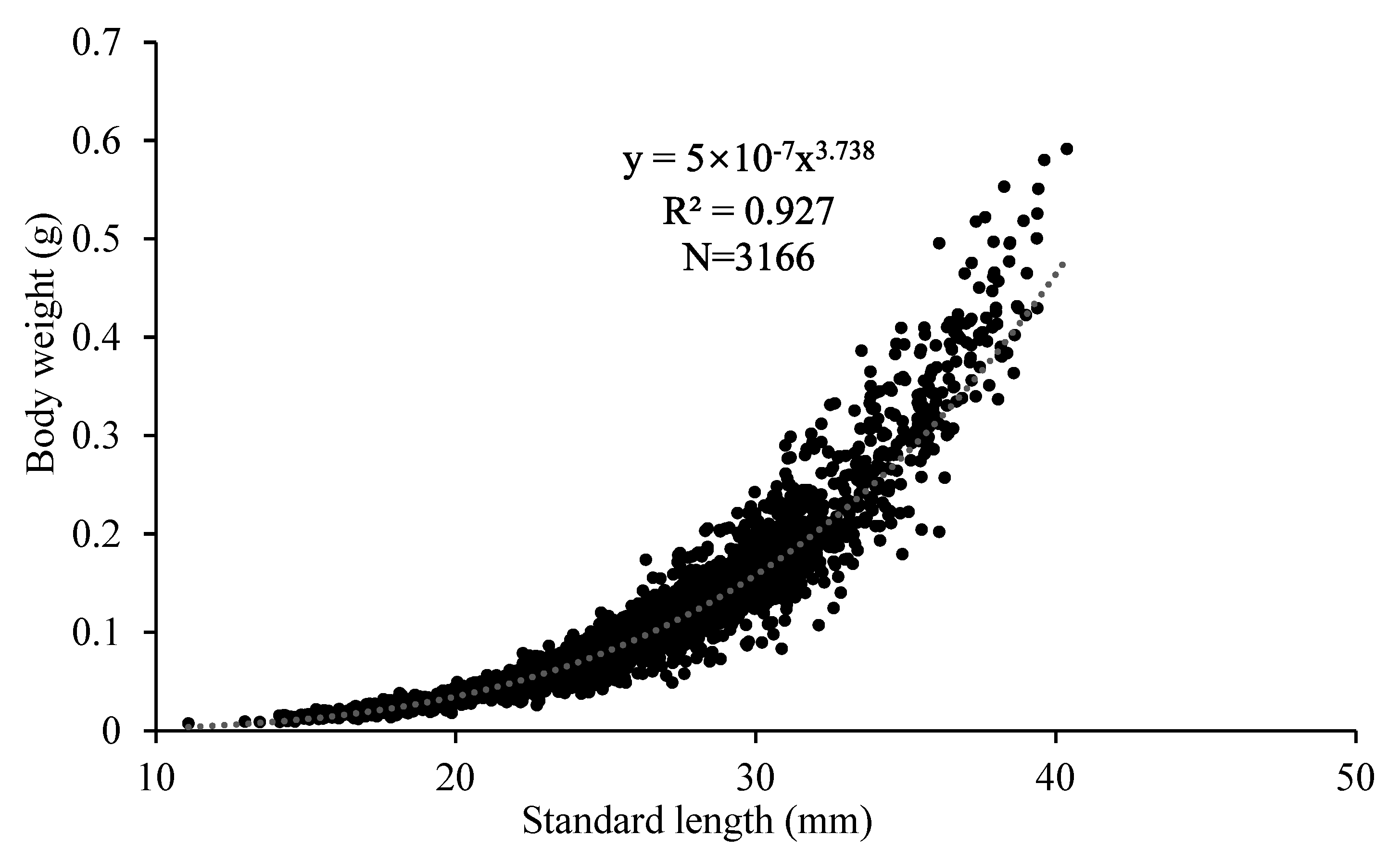
3.2. Length-Weight Relationship
3.3. Growth Function
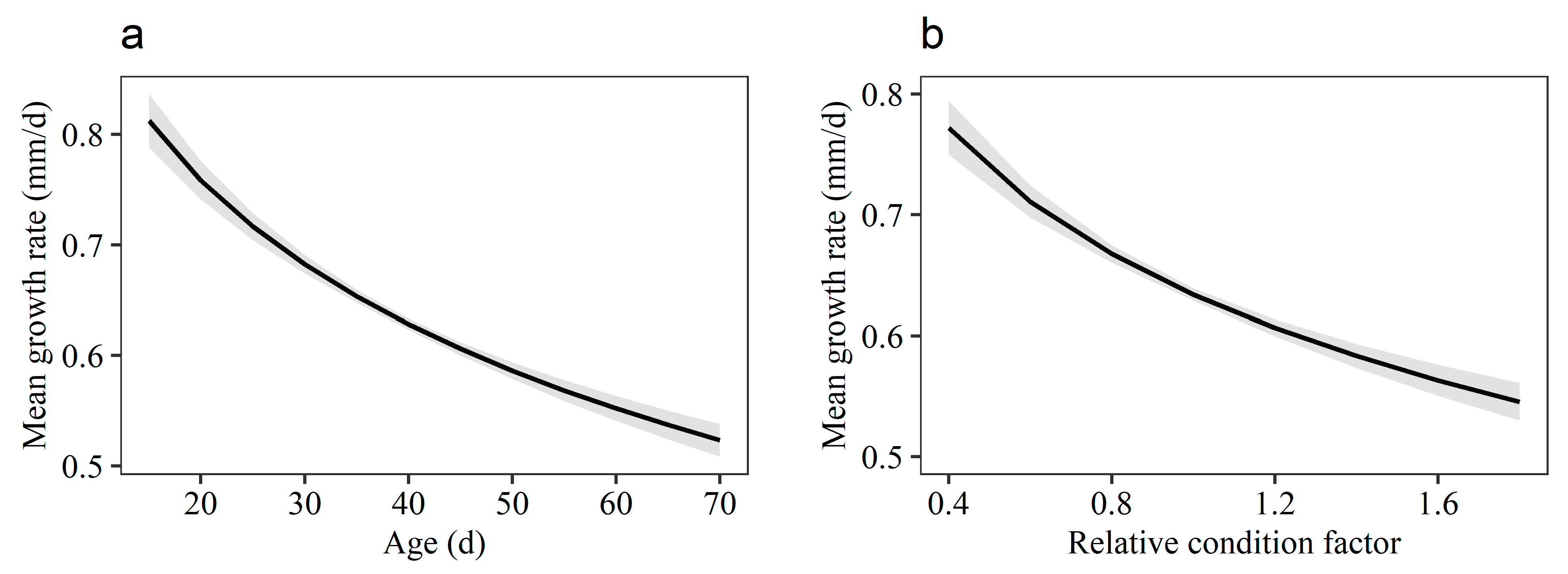
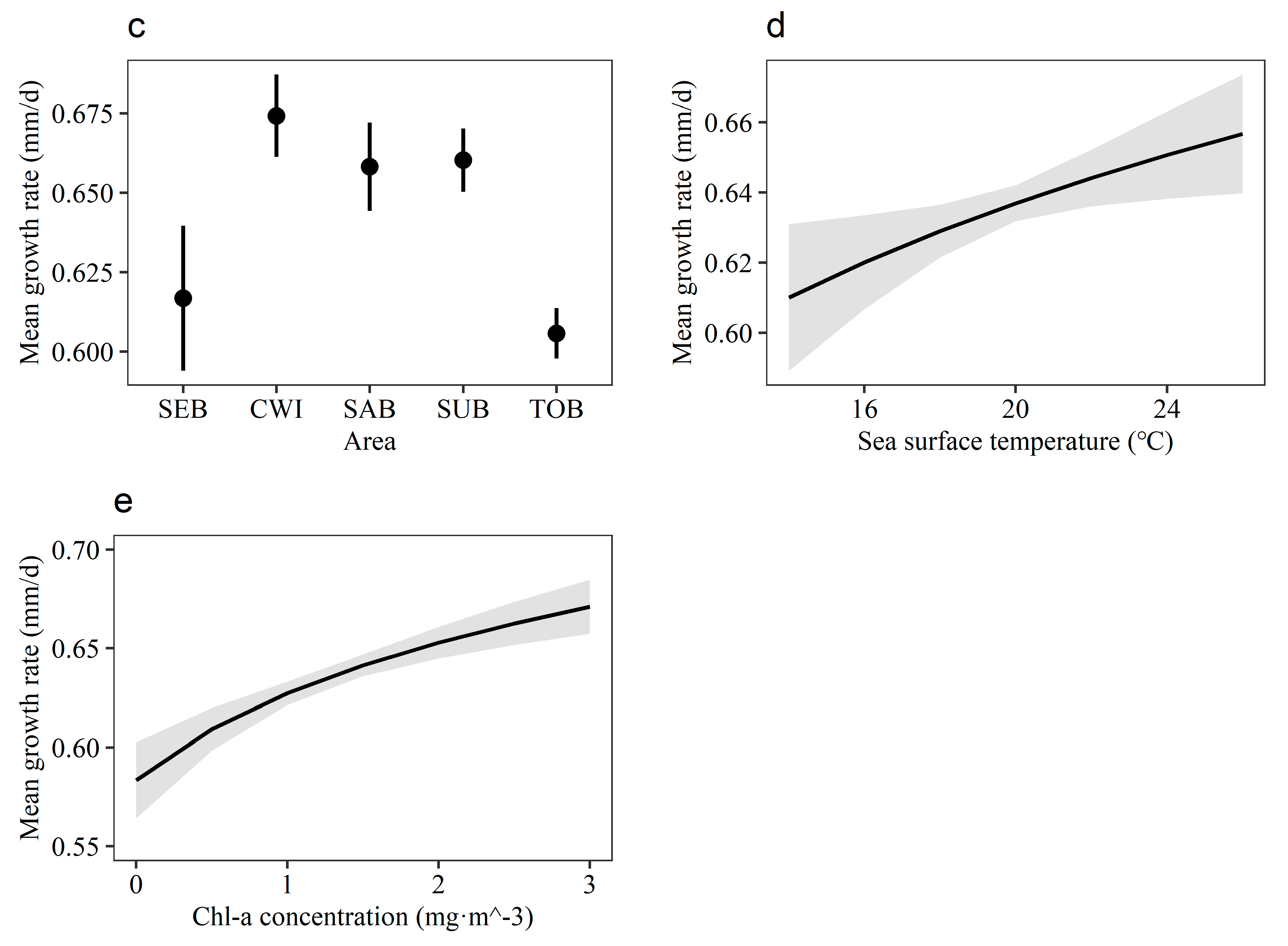
3.4. Growth Rate Variation among Individuals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asch, R.G.; Stock, C.A.; Sarmiento, J.L. Climate change impacts on mismatches between phytoplankton blooms and fish spawning phenology. Glob. Chang. Biol. 2019, 25, 2544–2559. [Google Scholar] [CrossRef] [PubMed]
- Cushing, D. Plankton Production and Year-class Strength in Fish Populations: An Update of the Match/Mismatch Hypothesis. In Advances in Marine Biology; Elsevier Applied Science Publishers Ltd.: London, UK, 1990; Volume 26, pp. 249–293. [Google Scholar] [CrossRef]
- Houde, E.D. Emerging from Hjort's Shadow. J. Northwest Atl. Fish. Sci. 2008, 41, 53–70. [Google Scholar] [CrossRef]
- Takasuka, A.; Sakai, A.; Aoki, I. Dynamics of growth-based survival mechanisms in Japanese anchovy (Engraulis japonicus) larvae. Can. J. Fish. Aquat. Sci. 2017, 74, 812–823. [Google Scholar] [CrossRef] [Green Version]
- Morrongiello, J.R.; Walsh, C.T.; Gray, C.A.; Stocks, J.R.; Crook, D.A. Environmental change drives long-term recruitment and growth variation in an estuarine fish. Glob. Chang. Biol. 2014, 20, 1844–1860. [Google Scholar] [CrossRef]
- Cowan, J.J.; Rose, K.; Devries, D. Is density-dependent growth in young-of-the-year fishes a question of critical weight? Rev. Fish Biol. Fish. 2000, 10, 61–89. [Google Scholar] [CrossRef]
- Whitten, A.R.; Klaer, N.L.; Tuck, G.N.; Day, R.W. Accounting for cohort-specific variable growth in fisheries stock assessments: A case study from south-eastern Australia. Fish. Res. 2013, 142, 27–36. [Google Scholar] [CrossRef]
- Lluch-Belda, D.; Crawford, R.J.M.; Kawasaki, T.; MacCall, A.D.; Parrish, R.H.; Schwartzlose, R.A.; Smith, P.E. World-wide fluctuations of sardine and anchovy stocks: The regime problem. South Afr. J. Mar. Sci. 1989, 8, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Schwartzlose, R.A.; Alheit, J.; Bakun, A.; Baumgartner, T.R.; Cloete, R.; Crawford, R.J.M.; Fletcher, W.J.; Green-Ruiz, Y.; Hagen, E.; Kawasaki, T.; et al. Worldwide large-scale fluctuations of sardine and anchovy populations. South Afr. J. Mar. Sci. 1999, 21, 289–347. [Google Scholar] [CrossRef]
- Takahashi, M.; Watanabe, Y. Staging larval and early juvenile Japanese anchovy based on the degree of guanine deposition. J. Fish Biol. 2004, 64, 262–267. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Zhang, Y.; Liu, Q.; Liu, F.; Li, D.; Zhang, H. Climate-Driven Synchrony in Anchovy Fluctuations: A Pacific-Wide Comparison. Fishes 2022, 7, 193. [Google Scholar] [CrossRef]
- Boldt, J.L.; Thompson, M.; Rooper, C.N.; Hay, D.E.; Schweigert, J.F.; Quinn, T.J., II; Cleary, J.S.; Neville, C.M. Bottom-up and top-down control of small pelagic forage fish: Factors affecting age-0 herring in the Strait of Georgia, British Columbia. Mar. Ecol. Prog. Ser. 2019, 617, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, M.; Fujita, T.; Yamamoto, M.; Tadokoro, K.; Okazaki, Y.; Nakamura, M.; Takahashi, M.; Kono, N.; Matsubara, T.; Abo, K.; et al. Bottom-up processes drive reproductive success of Japanese anchovy in an oligotrophic sea: A case study in the central Seto Inland Sea, Japan. Prog. Oceanogr. 2022, 206, 102860. [Google Scholar] [CrossRef]
- Kinoshita, J.; Yasuda, T.; Watanabe, C.; Kamimura, Y. Stock assessment and evaluation for Japanese anchovy Pacific stock (fiscal year 2021). In Marine Fisheries Stock Assessment and Evaluation for Japanese Waters; Japan Fisheries Agency and Japan Fisheries Research and Education Agency: Tokyo, Japan, 2022; p. 50. Available online: https://abchan.fra.go.jp/digests2021/index.html (accessed on 17 October 2022). (In Japanese)
- Tsuruta, Y. Internal regulation of reproduction in Japanese anchovy (Engraulis japonica) as related to population fluctuation. Can. Spec. Pub. Fish. Aquat. Sci. 1989, 108, 111–119. [Google Scholar]
- Yatsu, A. Review of population dynamics and management of small pelagic fishes around the Japanese Archipelago. Fish. Sci. 2019, 85, 611–639. [Google Scholar] [CrossRef] [Green Version]
- Funakoshi, S. Relationship between stock levels and the population structure of the Japanese anchovy. Mar. Behav. Physiol. 1992, 21, 1–84. [Google Scholar] [CrossRef]
- Funamoto, T.; Aoki, I.; Wada, Y. Reproductive characteristics of Japanese anchovy, Engraulis japonicus, in two bays of Japan. Fish. Res. 2004, 70, 71–81. [Google Scholar] [CrossRef]
- Takasuka, A.; Oozeki, Y.; Kubota, H.; Tsuruta, Y.; Funamoto, T. Temperature impacts on reproductive parameters for Japanese anchovy: Comparison between inshore and offshore waters. Fish. Res. 2005, 76, 475–482. [Google Scholar] [CrossRef]
- Hayashi, A.; Goto, T.; Takahashi, M.; Watanabe, Y. How Japanese anchovy spawn in northern waters: Start with surface warming and end with day length shortening. Ichthyol. Res. 2019, 66, 79–87. [Google Scholar] [CrossRef]
- Hayasi, S. A note on the biology and fishery of the Japanese anchovy Engraulis japonica (Houttuyn). Rep. Calif. Coop. Ocean. Fish. Investig. 1967, 11, 44–57. Available online: http://www.calcofi.com/publications/calcofireports/v11/Vol_11_Hayasi.pdf (accessed on 11 November 2022).
- Yamamoto, K.; Saito, M.; Yamashita, Y. Relationships between the daily growth rate of Japanese anchovy Engraulis japonicus larvae and environmental factors in Osaka Bay, Seto Inland Sea, Japan. Fish. Sci. 2018, 84, 373–383. [Google Scholar] [CrossRef]
- Takasuka, A.; Aoki, I. Environmental determinants of growth rates for larval Japanese anchovy Engraulis japonicus in different waters. Fish. Oceanogr. 2006, 15, 139–149. [Google Scholar] [CrossRef]
- Tsujino, K. Daily growth of Japanese anchovy larvae Engraulis japonica in Osaka Bay. Bull. Osaka Prefect. Fish. Exp. Sta. 2001, 13, 11–18. (In Japanese) [Google Scholar]
- Yasue, N.; Takasuka, A. Seasonal variability in growth of larval Japanese anchovy Engraulis japonicus driven by fluctuations in sea temperature in the Kii Channel, Japan. J. Fish Biol. 2009, 74, 2250–2268. [Google Scholar] [CrossRef] [PubMed]
- Takasuka, A.; Oozeki, Y.; Aoki, I.; Kimura, R.; Kubota, H.; Sugisaki, H.; Akamine, T. Growth effect on the otolith and somatic size relationship in Japanese anchovy and sardine larvae. Fish. Sci. 2008, 74, 308–313. [Google Scholar] [CrossRef]
- Tanner, S.E.; Vieira, A.R.; Vasconcelos, R.P.; Dores, S.; Azevedo, M.; Cabral, H.N.; Morrongiello, J.R. Regional climate, primary productivity and fish biomass drive growth variation and population resilience in a small pelagic fish. Ecol. Indic. 2019, 103, 530–541. [Google Scholar] [CrossRef]
- Froese, R.; Thorson, J.T.; Reyes, R.B. A Bayesian approach for estimating length-weight relationships in fishes. J. Appl. Ichthyol. 2013, 30, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.-B.; Zhu, H.-C.; Wang, Y.-L.; Zhang, Y.-Z.; Lu, Z.-H.; Cui, G.-C. Heterogeneity of fork length-weight relationship for juvenile Engraulis japonicus based on linear mixed-effects models. J. Appl. Ecol. 2021, 32, 4532–4538. (In Chinese) [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Koushlesh, S.K.; Sinha, A.; Kumari, K.; Borah, S.; Chanu, T.N.; Baitha, R.; Das, S.K.; Gogoi, P.; Sharma, S.K.; Ramteke, M.H.; et al. Length-weight relationship and relative condition factor of five indigenous fish species from Torsa River, West Bengal, India. J. Appl. Ichthyol. 2017, 34, 169–171. [Google Scholar] [CrossRef]
- Jisr, N.; Younes, G.; Sukhn, C.; El-Dakdouki, M.H. Length-weight relationships and relative condition factor of fish inhabiting the marine area of the Eastern Mediterranean city, Tripoli-Lebanon. Egypt. J. Aquat. Res. 2018, 44, 299–305. [Google Scholar] [CrossRef]
- Le Cren, E.D. The Length-Weight Relationship and Seasonal Cycle in Gonad Weight and Condition in the Perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Kodama, M.; Diamante, R.A.; Salayo, N.D.; Castel, R.J.G.; Sumbing, J.G. Growth Performance and Condition Factor of Juvenile Milkfish (Chanos chanos) Cultured in a Marine Pen in Relation to Body Size and Temperature. Jpn. Agric. Res. Quarterly: JARQ 2021, 55, 191–200. [Google Scholar] [CrossRef]
- Allan, B.J.M.; Browman, H.I.; Shema, S.; Skiftesvik, A.-B.; Folkvord, A.; Durif, C.M.F.; Kjesbu, O.S. Increasing temperature and prey availability affect the growth and swimming kinematics of Atlantic herring (Clupea harengus) larvae. J. Plankton Res. 2022, 44, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Raab, K.; Llope, M.; Nagelkerke, L.A.; Rijnsdorp, A.D.; Teal, L.R.; Licandro, P.; Ruardij, P.; Dickey-Collas, M. Influence of temperature and food availability on juvenile European anchovy Engraulis encrasicolus at its northern boundary. Mar. Ecol. Prog. Ser. 2013, 488, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Zenitani, H.; Kono, N.; Tsukamoto, Y.; Masuda, R. Effects of temperature, food availability, and body size on daily growth rate of Japanese anchovy Engraulis japonicus larvae in Hiuchi-nada. Fish. Sci. 2009, 75, 1177–1188. [Google Scholar] [CrossRef]
- Cermeño, P.; Uriarte, A.; Morales-Nin, B.; Cotano, U.; Álvarez, P. Setting up interpretation criteria for ageing juvenile european anchovy otoliths. Sci. Mar. 2008, 72, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.-D.; Song, M.-H.; Lee, T.-W.; McFarlane, G.A.; King, J.R. Growth of larval Pacific anchovy Engraulis japonicus in the Yellow Sea as indicated by otolith microstructure analysis. J. Fish Biol. 2006, 69, 1756–1769. [Google Scholar] [CrossRef]
- Takahashi, M.; Watanabe, Y.; Kinoshita, T.; Watanabe, C. Growth of larval and early juvenile Japanese anchovy, Engraulis japonicus, in the Kuroshio-Oyashio transition region. Fish. Oceanogr. 2001, 10, 235–247. [Google Scholar] [CrossRef]
- Tsuji, S.; Aoyama, T. Daily growth increments in otoliths of Japanese anchovy larvae Engraulis japonica. Nippon Suisan Gakkaishi 1984, 50, 1105–1108. [Google Scholar] [CrossRef] [Green Version]
- Namiki, S.; Tanaka, H.; Katayama, S.; Funaki, O.; Aoki, I.; Oozeki, Y. Validation of daily increment formation in otoliths of immature and adult Japanese anchovy Engraulis japonicus. Fish. Sci. 2010, 76, 951–959. [Google Scholar] [CrossRef]
- Takasuka, A.; Aoki, I.; Mitani, I. Three synergistic growth-related mechanisms in the short-term survival of larval Japanese anchovy Engraulis japonicus in Sagami Bay. Mar. Ecol. Prog. Ser. 2004, 270, 217–228. [Google Scholar] [CrossRef]
- Chiu, T.; Chen, C. Growth and temporal variation of two Japanese anchovy cohorts during their recruitment to the East China Sea. Fish. Res. 2001, 53, 1–15. [Google Scholar] [CrossRef]
- Katsanevakis, S. Modelling fish growth: Model selection, multi-model inference and model selection uncertainty. Fish. Res. 2006, 81, 229–235. [Google Scholar] [CrossRef]
- Ong, J.J.L.; Rountrey, A.N.; Marriott, R.J.; Newman, S.J.; Meeuwig, J.J.; Meekan, M.G. Cross-continent comparisons reveal differing environmental drivers of growth of the coral reef fish, Lutjanus bohar. Coral Reefs 2016, 36, 195–206. [Google Scholar] [CrossRef]
- Chang, S.-K.; Chou, Y.-T.; Hoyle, S.D. Length-Weight Relationships and Otolith-Based Growth Curves for Brushtooth Lizardfish off Taiwan With Observations of Region and Aging–Material Effects on Global Growth Estimates. Front. Mar. Sci. 2022, 9, 921594. [Google Scholar] [CrossRef]
- Fukuhara, O.; Takao, K. Growth and larval behaviour of Engraulis japonica in captivity. J. Appl. Ichthyol. 1988, 4, 158–167. [Google Scholar] [CrossRef]
- Doubleday, Z.A.; Izzo, C.; Haddy, J.A.; Lyle, J.M.; Ye, Q.; Gillanders, B.M. Long-term patterns in estuarine fish growth across two climatically divergent regions. Oecologia 2015, 179, 1079–1090. [Google Scholar] [CrossRef]
- Masuda, R. Ontogeny of swimming speed, schooling behaviour and jellyfish avoidance by Japanese anchovy Engraulis japonicus. J. Fish Biol. 2011, 78, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Oozeki, Y.; Takasuka, A.; Kubota, H.; Barange, M. Characterizing spawning habitats of Japanese sardine, Sardinops melanostictus, Japanese anchovy, Engraulis japonicus, and Pacific round herring, Etrumeus teres, in the Northwestern Pacific. Calif. Coop. Ocean. Fish. Investig. Rep. 2007, 48, 191. [Google Scholar]
- Okata, A. Ecological studies on the biological production of young amberfish community in the Sendai Bay. II. Relationships between food chains and fish fauna. Nippon Suisan Gakkaishi 1976, 42, 29–44. [Google Scholar] [CrossRef]
- Bailey, K.; Houde, E. Predation on Eggs and Larvae of Marine Fishes and the Recruitment Problem. In Advances in Marine Biology; Blaxter, J.H.S., Southward, A.J., Eds.; Academic Press Inc.: New York, NY, USA, 1969; Volume 25, pp. 1–83. [Google Scholar] [CrossRef]
- Takasuka, A.; Aoki, I.; Mitani, I. Evidence of growth-selective predation on larval Japanese anchovy Engraulis japonicus in Sagami Bay. Mar. Ecol. Prog. Ser. 2003, 252, 223–238. [Google Scholar] [CrossRef]
- Takasuka, A.; Aoki, I.; Oozeki, Y. Predator-specific growth-selective predation on larval Japanese anchovy Engraulis japonicus. Mar. Ecol. Prog. Ser. 2007, 350, 99–107. [Google Scholar] [CrossRef]
- Fujita, T.; Yamamoto, M.; Kono, N.; Tomiyama, T.; Sugimatsu, K.; Yoneda, M. Temporal variations in hatch date and early survival of Japanese anchovy ( Engraulis japonicus ) in response to environmental factors in the central Seto Inland Sea, Japan. Fish. Oceanogr. 2021, 30, 527–541. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication. Available online: https://www.fishbase.org (accessed on 22 August 2022).
- Kimmerer, W.; Avent, S.R.; Bollens, S.M.; Feyrer, F.; Grimaldo, L.F.; Moyle, P.B.; Nobriga, M.; Visintainer, T. Variability in Length–Weight Relationships Used to Estimate Biomass of Estuarine Fish from Survey Data. Trans. Am. Fish. Soc. 2005, 134, 481–495. [Google Scholar] [CrossRef]
- MacFarlane, R.B.; Norton, E.C. Physiological ecology of juvenile Chinook salmon (Oncorhynchus tshawytscha) at the southern end of their distribution, the San Francisco Estuary and Gulf of the Farallones, California. Fish. Bull. 2002, 100, 244–257. [Google Scholar]
- Flinn, S.A.; Midway, S.R. Trends in Growth Modeling in Fisheries Science. Fishes 2021, 6, 1. [Google Scholar] [CrossRef]
- Schnute, J. A Versatile Growth Model with Statistically Stable Parameters. Can. J. Fish. Aquat. Sci. 1981, 38, 1128–1140. [Google Scholar] [CrossRef]
- Plaza, G.; Cerna, F.; Landaeta, M.F.; Hernández, A.; Contreras, J.E. Daily growth patterns and age-at-recruitment of the anchoveta Engraulis ringens as indicated by a multi-annual analysis of otolith microstructure across developmental stages. J. Fish Biol. 2018, 93, 370–381. [Google Scholar] [CrossRef]
- Dulčić, J. Growth of anchovy, Engraulis encrasicolus (L.), larvae in the Northern Adriatic Sea. Fish. Res. 1997, 31, 189–195. [Google Scholar] [CrossRef]
- Basilone, G.; Ferreri, R.; Mangano, S.; Pulizzi, M.; Gargano, A.; Barra, M.; Mazzola, S.; Fontana, I.; Giacalone, G.; Genovese, S.; et al. Effects of habitat conditions at hatching time on growth history of offspring European anchovy, Engraulis encrasicolus, in the Central Mediterranean Sea. Hydrobiologia 2018, 821, 99–111. [Google Scholar] [CrossRef]
- Hernandez, E.H.; Castro, L.R. Larval growth of the anchoveta Engraulis ringens during the winter spawning season off central Chile. Fish. Bull. 2000, 98, 704. [Google Scholar]
- Tabatabai, M.; Williams, D.K.; Bursac, Z. Hyperbolastic growth models: Theory and application. Theor. Biol. Med. Model. 2005, 2, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shima, J.S.; Osenberg, C.W.; Noonburg, E.G.; Alonzo, S.H.; Swearer, S.E. Lunar rhythms in growth of larval fish. Proc. R. Soc. B: Biol. Sci. 2021, 288, 20202609. [Google Scholar] [CrossRef] [PubMed]
- Cordoleani, F.; Holmes, E.; Bell-Tilcock, M.; Johnson, R.C.; Jeffres, C. Variability in foodscapes and fish growth across a habitat mosaic: Implications for management and ecosystem restoration. Ecol. Indic. 2022, 136, 108681. [Google Scholar] [CrossRef]
- Nishikawa, H.; Itoh, S.; Yasuda, I.; Komatsu, K. Overlap between suitable nursery grounds for Japanese anchovy (Engraulis japonicus) and Japanese sardine (Sardinops melanostictus) larvae. Aquac. Fish Fish. 2022, 2, 179–188. [Google Scholar] [CrossRef]
- Mitani, I. The biological studies on the larvae of Japanese anchovy, Engraulis japonica Houttuyn, in Sagami Bay. Kanagawa Pref. Fish. Exp. Sta. Rep. 1990, 5, 1–140. (In Japanese) [Google Scholar]
- Aoki, I.; Miyashita, K. Dispersal of larvae and juveniles of Japanese anchovy Engraulis japonicus in the Kuroshio Extension and Kuroshio–Oyashio transition regions, Western North Pacific Ocean. Fish. Res. 2000, 49, 155–164. [Google Scholar] [CrossRef]
- Werner, E.E.; Anholt, B.R. Ecological Consequences of the Trade-Off between Growth and Mortality Rates Mediated by Foraging Activity. Am. Nat. 1993, 142, 242–272. [Google Scholar] [CrossRef]
- Biro, P.A.; Post, J.R. Rapid depletion of genotypes with fast growth and bold personality traits from harvested fish populations. Proc. Natl. Acad. Sci. USA 2008, 105, 2919–2922. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, M.; Kitano, H.; Tanaka, H.; Kawamura, K.; Selvaraj, S.; Ohshimo, S.; Matsuyama, M.; Shimizu, A. Temperature- and income resource availability-mediated variation in reproductive investment in a multiple-batch-spawning Japanese anchovy. Mar. Ecol. Prog. Ser. 2014, 516, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.M.; Buehrens, T.W.; Fisher, J.L.; Wilson, K.L.; Moore, J.W. Phenological mismatch, carryover effects, and marine survival in a wild steelhead trout Oncorhynchus mykiss population. Prog. Oceanogr. 2021, 193, 102533. [Google Scholar] [CrossRef]
- Reznick, D.; Lindbeck, E.; Bryga, H. Slower Growth Results in Larger Otoliths: An Experimental Test with Guppies (Poecilia reticulata). Can. J. Fish. Aquat. Sci. 1989, 46, 108–112. [Google Scholar] [CrossRef]
- Macdonald, P.; Angus, C.H.; Marshall, C.T. Spatial variation in life history characteristics of common megrim (Lepidorhombus whiffiagonis) on the Northern Shelf. J. Sea Res. 2013, 75, 62–68. [Google Scholar] [CrossRef]
- Barrios, A.; Ernande, B.; Mahé, K.; Trenkel, V.; Rochet, M.-J. Utility of mixed effects models to inform the stock structure of whiting in the Northeast Atlantic Ocean. Fish. Res. 2017, 190, 132–139. [Google Scholar] [CrossRef] [Green Version]
- McBride, R.S. The Continuing Role of Life History Parameters to Identify Stock Structure. In Stock Identification Methods, 2nd ed.; Cadrin, S.X., Kerr, L.A., Mariani, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 77–107. [Google Scholar] [CrossRef]
- van Denderen, D.; Gislason, H.; van den Heuvel, J.; Andersen, K.H. Global analysis of fish growth rates shows weaker responses to temperature than metabolic predictions. Glob. Ecol. Biogeogr. 2020, 29, 2203–2213. [Google Scholar] [CrossRef]
- Kuwahara, A. Diurnal changes in vertical distributions of anchovy eggs and larvae in the western Wakasa Bay. Nippon Suisan Gakkaishi 1984, 50, 1285–1292. [Google Scholar] [CrossRef]
- Takahashi, M.; Watanabe, Y. Effects of temperature and food availability on growth rate during late larval stage of Japanese anchovy (Engraulis japonicus) in the Kuroshio-Oyashio transition region. Fish. Oceanogr. 2005, 14, 223–235. [Google Scholar] [CrossRef]
- Zenitani, H.; Kono, N.; Tsukamoto, Y. Relationship between daily survival rates of larval Japanese anchovy (Engraulis japonicus) and concentrations of copepod nauplii in the Seto Inland Sea, Japan. Fish. Oceanogr. 2007, 16, 473–478. [Google Scholar] [CrossRef]
- Yasue, N.; Doiuchi, R.; Yoshimoto, Y.; Takeuchi, T. Diet of late larval Japanese anchovy Engraulis japonicus in the Kii Channel, Japan. Fish. Sci. 2009, 76, 63. [Google Scholar] [CrossRef]
- Houde, E.D. Mortality. In Fishery Science: The Unique Contributions of Early Life Stages; Fuiman, L.A., Werner, R.G., Eds.; Blackwell Publishing: Oxford, UK, 2002; pp. 64–87. [Google Scholar]
- Garrido, S.; Saiz, E.; Peters, J.; Ré, P.; Alvarez, P.; Cotano, U.; Herrero, D.L.; de Murguía, A.M.; Irigoien, X. Effect of food type and concentration on growth and fatty acid composition of early larvae of the anchovy (Engraulis encrasicolus) reared under laboratory conditions. J. Exp. Mar. Biol. Ecol. 2012, 434–435, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Huot, Y.; Babin, M.; Bruyant, F.; Grob, C.; Twardowski, M.S.; Claustre, H. Does chlorophyll a provide the best index of phytoplankton biomass for primary productivity studies? Biogeosciences 2007, 4, 707–745. [Google Scholar] [CrossRef] [Green Version]
- Wernand, M.R.; Van der Woerd, H.J.; Gieskes, W.W. Trends in Ocean Colour and Chlorophyll Concentration from 1889 to 2000, Worldwide. PLoS ONE 2013, 8, e63766. [Google Scholar] [CrossRef] [PubMed]
- Cowan, J.H., Jr.; Rose, K.A.; Houde, E.D.; Wang, S.-B.; Young, J. Modeling effects of increased larval mortality on bay anchovy population dynamics in the mesohaline Chesapeake Bay: Evidence for compensatory reserve. Mar. Ecol. Prog. Ser. 1999, 185, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Boëns, A.; Grellier, P.; Lebigre, C.; Petitgas, P. Determinants of growth and selective mortality in anchovy and sardine in the Bay of Biscay. Fish. Res. 2021, 239, 105947. [Google Scholar] [CrossRef]
- Ekau, W.; Auel, H.; Pörtner, H.-O.; Gilbert, D. Impacts of hypoxia on the structure and processes in pelagic communities (zooplankton, macro-invertebrates and fish). Biogeosciences 2010, 7, 1669–1699. [Google Scholar] [CrossRef] [Green Version]
- Waldron, M.; Armstrong, M.J.; Prosch, R.M. Aspects of the variability in growth of juvenile anchovyEngraulis capensisin the southern Benguela system. South Afr. J. Mar. Sci. 1989, 8, 9–19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Area | n | Range of Age (d) | Range of SL (mm) | Back-Calculated Spawning Season | Back-Calculated Spawning SST (°C) |
|---|---|---|---|---|---|
| SEB | 92 | 23–67 | 18.11–32.91 | May–September | 12.2–27.1 |
| CWI | 228 | 22–50 | 17.82–33.80 | April–October | 12.7–25.8 |
| SAB | 146 | 21–44 | 18.13–32.49 | February–July | 15.7–23.4 |
| SUB | 293 | 23–54 | 16.21–39.38 | February–May | 15.8–18.2 |
| TOB | 599 | 17–64 | 12.96–36.29 | January–May | 16.9–20.8 |
| Model | R2 | AIC | ∆AIC | ||||
|---|---|---|---|---|---|---|---|
| Gompertz | 34.23 | 0.059 | 14.26 | 0.6094 | 2569.19 | 0 | |
| Von Bertalanffy | 35.87 | 0.042 | 5.22 | 0.6088 | 2569.29 | 0.10 | |
| Logistic | 33.28 | 0.076 | 19.38 | 0.6086 | 2570.03 | 0.84 | |
| Richards | 34.82 | 0.051 | 0.565 | 11.05 | 0.6086 | 2571.09 | 1.90 |
| Predictors | HD-Based Model | CD-Based Model | ||||
|---|---|---|---|---|---|---|
| Estimates | CI | p | Estimates | CI | p | |
| Intercept | −0.49 | (−0.53, −0.46) | <0.001 | −0.41 | (−0.51, −0.32) | <0.001 |
| Age | −0.29 | (−0.33, −0.25) | <0.001 | −0.32 | (−0.40, −0.25) | <0.001 |
| Kn | −0.24 | (−0.28, −0.21) | <0.001 | −0.21 | (−0.25, −0.17) | <0.001 |
| Area | <0.001 | <0.001 | ||||
| SST | 0.13 | (0.04, 0.22) | 0.004 | |||
| Chl-a | 0.10 | (0.07, 0.14) | <0.001 | |||
| Random effects | HD | CD | ||||
| ICC | 0.30 | 0.45 | ||||
| AICc | −2322.86 | −2596.44 | ||||
| Marginal R2 | 0.444 | 0.429 | ||||
| Conditional R2 | 0.611 | 0.688 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Wu, R.; Masuda, Y.; Takahashi, Y.; Okabe, K.; Koizumi, K.; Iida, A.; Katayama, S. Spawning Phenology and Early Growth of Japanese Anchovy (Engraulis japonicus) off the Pacific Coast of Japan. Fishes 2023, 8, 11. https://doi.org/10.3390/fishes8010011
Zhu Q, Wu R, Masuda Y, Takahashi Y, Okabe K, Koizumi K, Iida A, Katayama S. Spawning Phenology and Early Growth of Japanese Anchovy (Engraulis japonicus) off the Pacific Coast of Japan. Fishes. 2023; 8(1):11. https://doi.org/10.3390/fishes8010011
Chicago/Turabian StyleZhu, Qinghuan, Rui Wu, Yoshio Masuda, Yutaro Takahashi, Kyu Okabe, Koji Koizumi, Atsuko Iida, and Satoshi Katayama. 2023. "Spawning Phenology and Early Growth of Japanese Anchovy (Engraulis japonicus) off the Pacific Coast of Japan" Fishes 8, no. 1: 11. https://doi.org/10.3390/fishes8010011