
A New Non-Obese Steatohepatitis Mouse Model with Cardiac Dysfunction Induced by Addition of Ethanol to a High-Fat/High-Cholesterol Diet

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mouse Model and Diets
2.2. Echocardiography Examination
2.3. Blood and Tissue Collection
2.4. Histopathological and Fluorescence Immunohistochemical Examinations
2.5. Real-Time Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
2.6. Blood Chemistry Tests
2.7. Determination of Hepatic TG and TCHO Levels
2.8. Analysis of the Blood Morning-Residual Ethanol
2.9. Statistical Analyses
3. Results
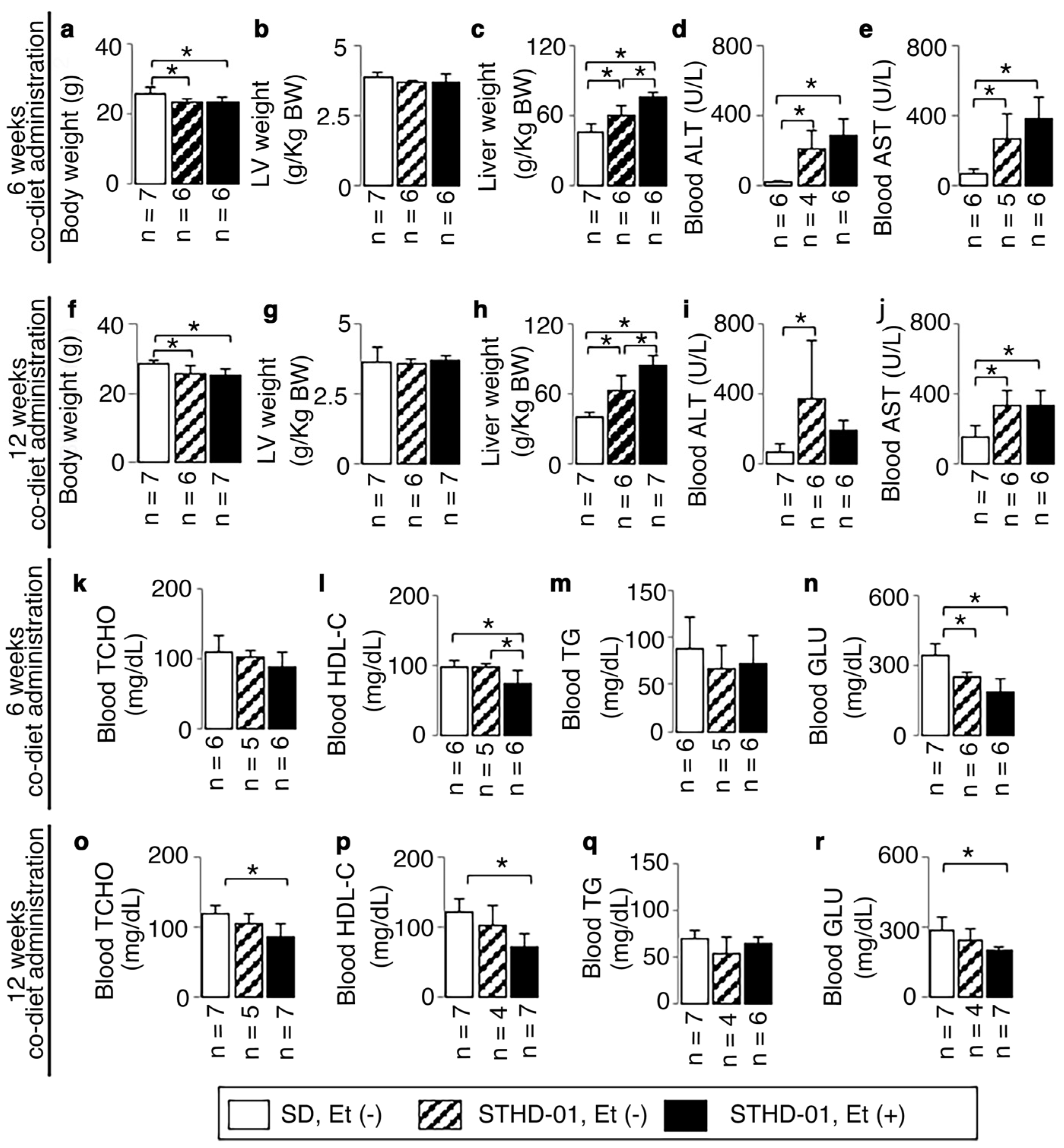
3.1. Animal Characteristics
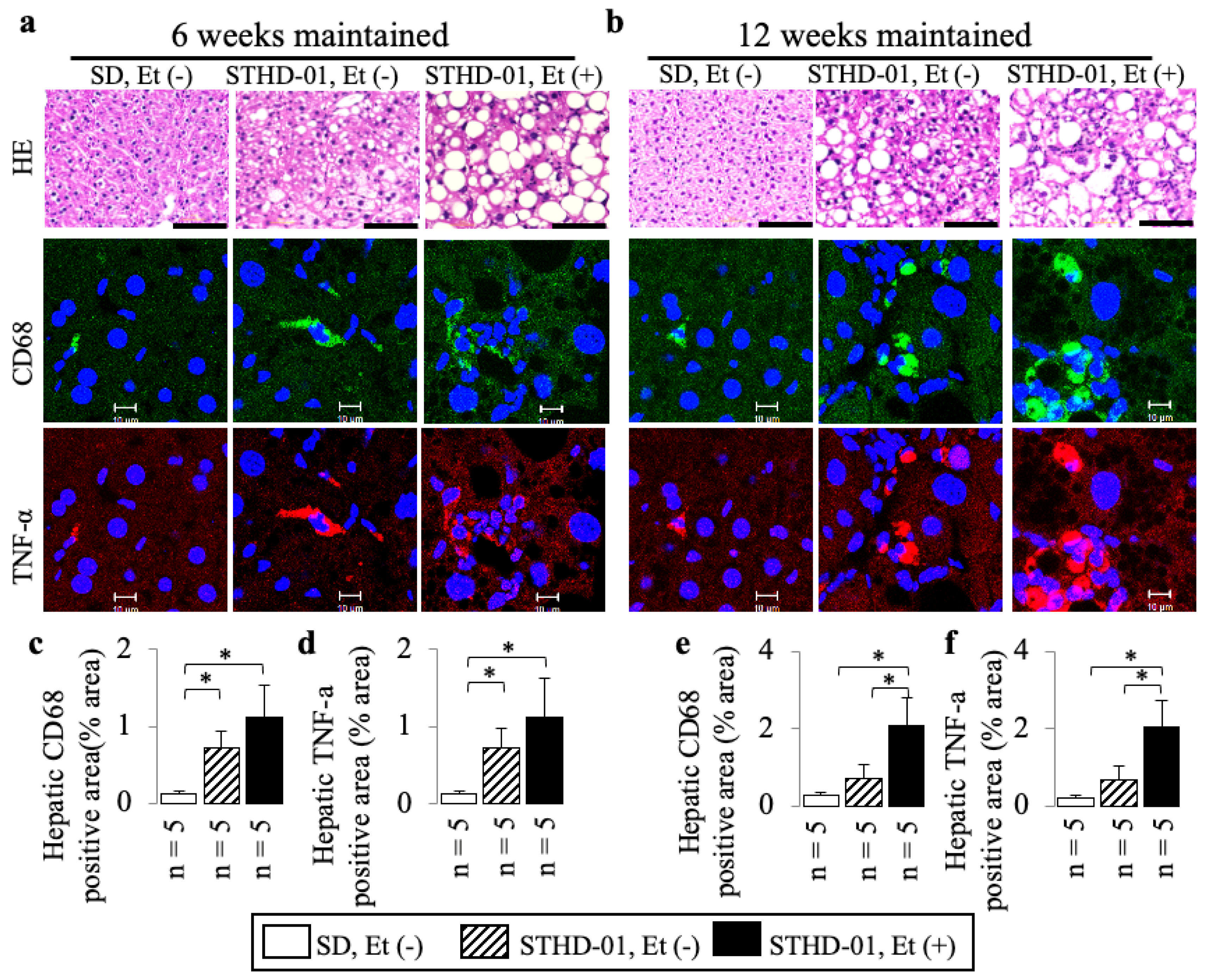
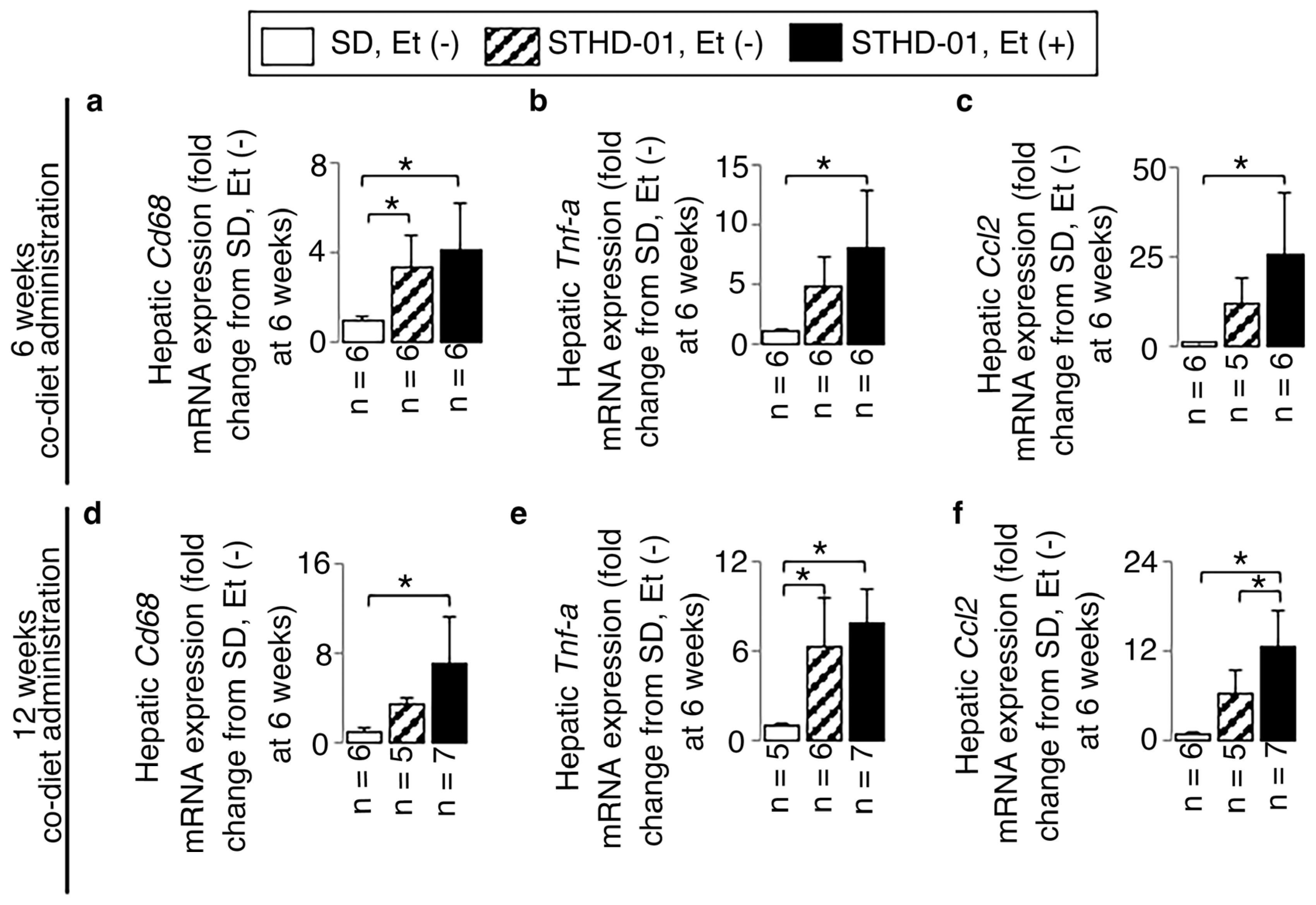
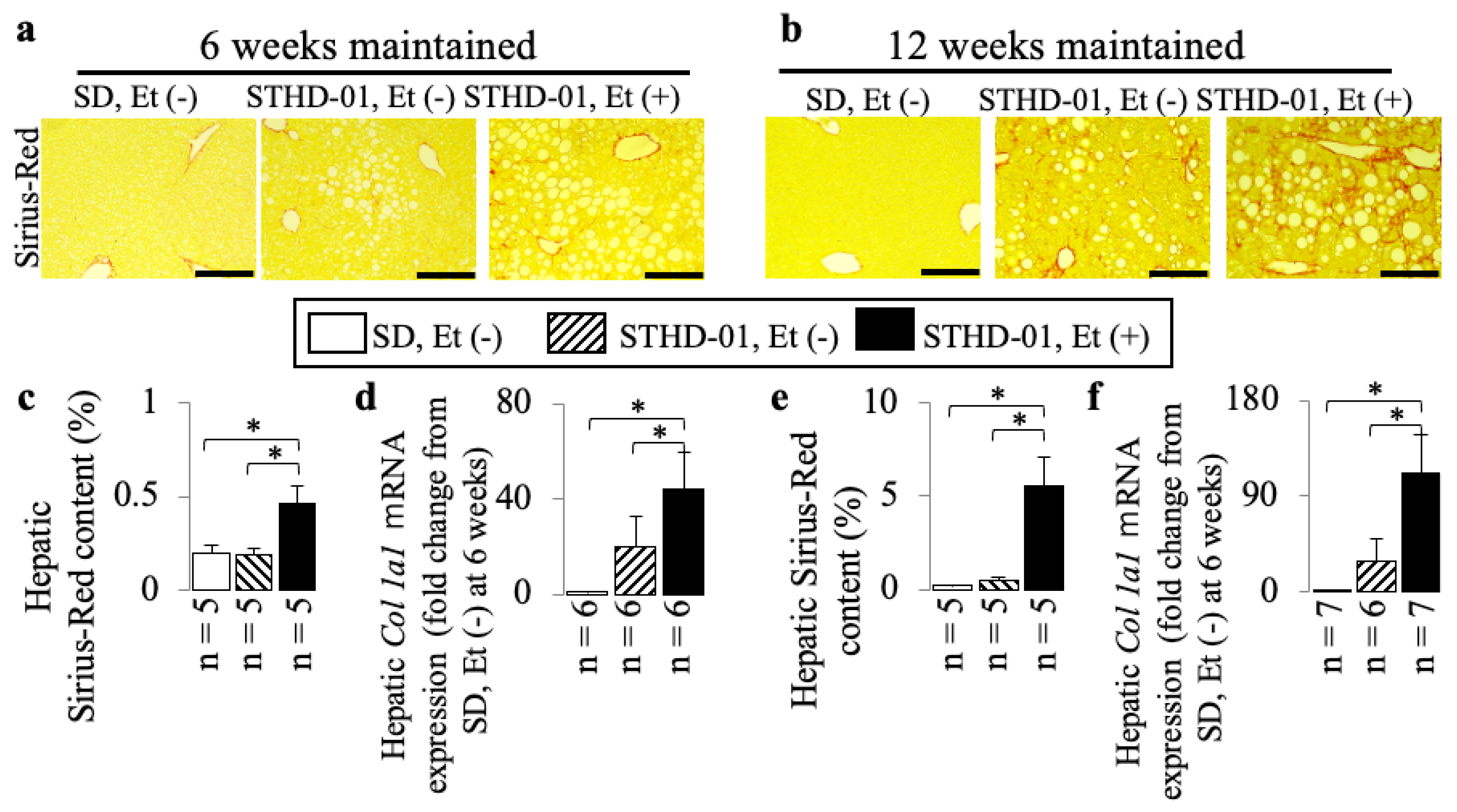
3.2. A Co-Diet of STHD-01 and Ethanol Produced Hepatic Steatosis, Inflammation, and Advanced Liver Fibrosis
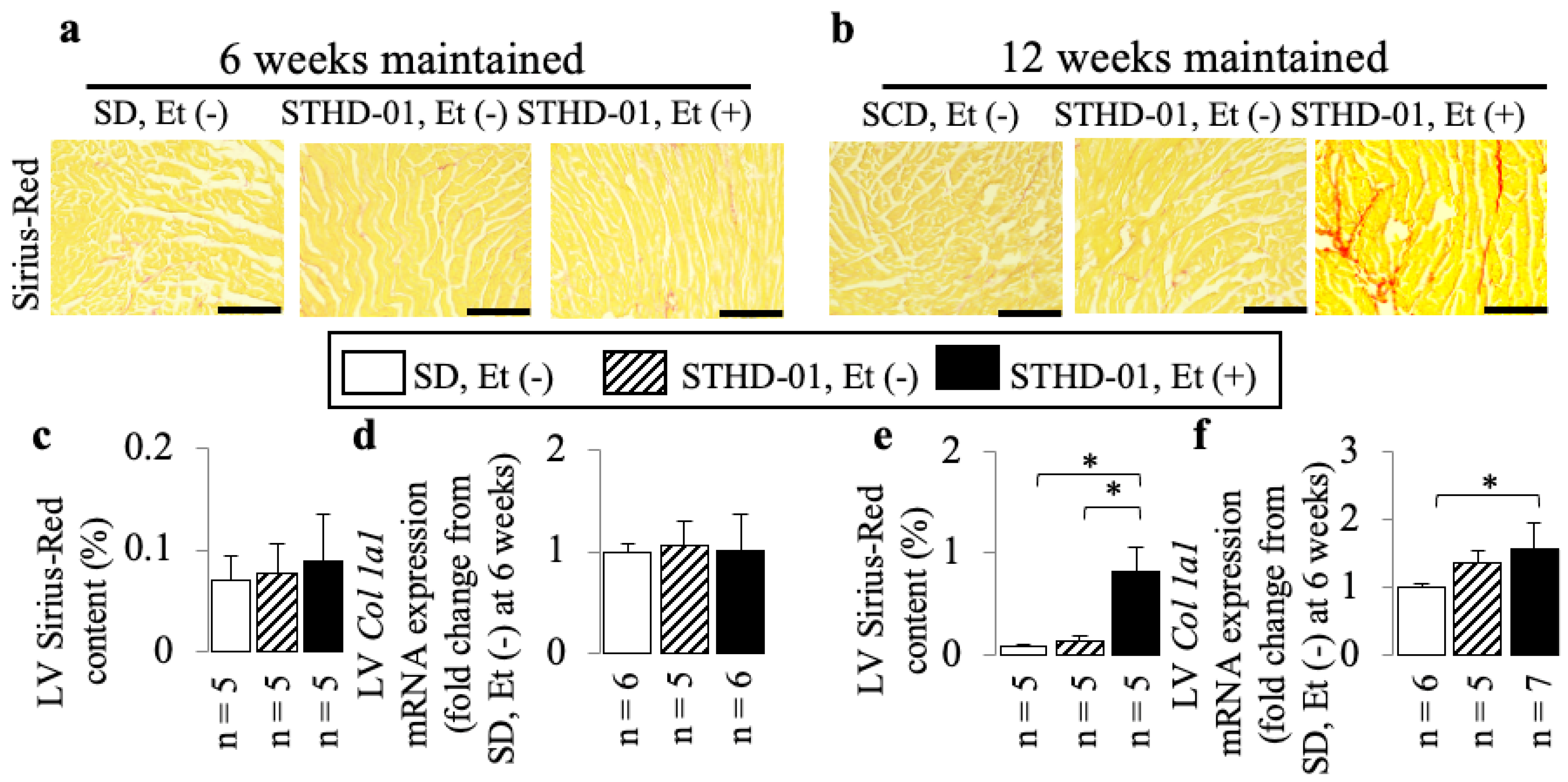
3.3. A Co-Diet of STHD-01 and Ethanol Caused Cardiac Dysfunction from 6 Weeks Onwards, and Cardiac Dilation with Myocardial Fibrosis at 12 Weeks of Administration
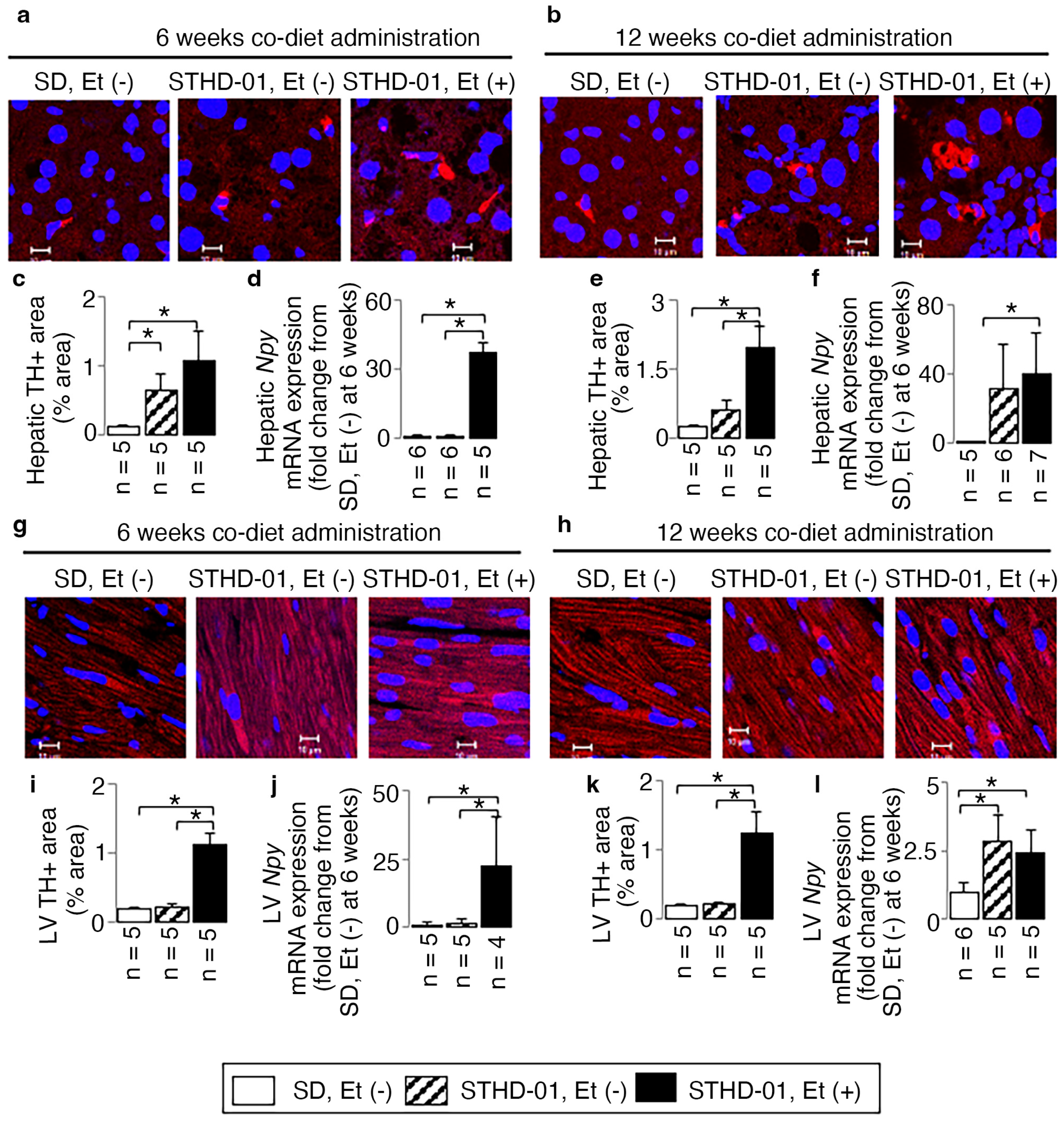
3.4. A Co-Diet of Ethanol and STHD-01 Induced Features of Hepatic and LV Myocardial Sympathetic Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Kumar, M.; Eslam, M.; George, J.; Al Mahtab, M.; Akbar, S.M.F.; Jia, J.; Tian, Q.; Aggarwal, R.; Muljono, D.H.; et al. Liver diseases in the Asia-Pacific region: A Lancet Gastroenterology & hepatology Commission. Lancet Gastroenterol. Hepatol. 2020, 5, 167–228. [Google Scholar]
- Ahadi, M.; Molooghi, K.; Masoudifar, N.; Namdar, A.B.; Vossoughinia, H.; Farzanehfar, M. A review of non-alcoholic fatty liver disease in non-obese and lean individuals. J. Gastroenterol. Hepatol. 2021, 36, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Stepanova, M.; Negro, F.; Hallaji, S.; Younossi, Y.; Lam, B.; Srishord, M. Nonalcoholic fatty liver disease in lean individuals in the United States. Medicine 2012, 91, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.G.; Kim, S.U.; Wong, V.W. New trends on obesity and NAFLD in Asia. J. Hepatol. 2017, 67, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zou, B.; Yeo, Y.H.; Li, J.; Huang, D.Q.; Wu, Y.; Yang, H.; Liu, C.; Kam, L.Y.; Tan, X.X.E.; et al. Global prevalence, incidence, and outcomes of non-obese or lean non-alcoholic fatty liver disease: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2020, 5, 739–752. [Google Scholar] [CrossRef]
- Lu, F.B.; Zheng, K.I.; Rios, R.S.; Targher, G.; Byrne, C.D.; Zheng, M.H. Global Epidemiology of Lean Non-Alcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. J. Gastroenterol. Hepatol. 2020, 35, 2041–2050. [Google Scholar] [CrossRef]
- Leung, J.C.F.; Loong, T.C.W.; Wei, J.L.; Wong, G.L.H.; Chan, A.W.H.; Choi, P.C.L.; Shu, S.S.T.; Chim, A.M.L.; Chan, H.L.Y.; Wong, V.W.S. Histological severity and clinical outcomes of nonalcoholic fatty liver disease in nonobese patients. Hepatology 2017, 65, 54–64. [Google Scholar] [CrossRef]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Investig. 2005, 115, 1343–1351. [Google Scholar] [CrossRef]
- Bugianesi, E.; Gastaldelli, A.; Vanni, E.; Gambino, R.; Cassader, M.; Baldi, S.; Ponti, V.; Pagano, G.; Ferrannini, E.; Rizzetto, M. Insulin resistance in non-diabetic patients with non-alcoholic fatty liver disease: Sites and mechanisms. Diabetologia 2005, 48, 634–642. [Google Scholar] [CrossRef]
- Valbusa, F.; Agnoletti, D.; Scala, L.; Grillo, C.; Arduini, P.; Bonapace, S.; Calabria, S.; Scaturro, G.; Mantovani, A.; Zoppini, G.; et al. Non-alcoholic fatty liver disease and increased risk of all-cause mortality in elderly patients admitted for acute heart failure. Int. J. Cardiol. 2018, 265, 162–168. [Google Scholar] [CrossRef]
- Nucera, S.; Ruga, S.; Cardamone, A.; Coppoletta, A.R.; Guarnieri, L.; Zito, M.C.; Bosco, F.; Macrì, R.; Scarano, F.; Scicchitano, M.; et al. MAFLD progression contributes to altered thalamus metabolism and brain structure. Sci. Rep. 2022, 12, 1027. [Google Scholar] [CrossRef]
- Tsutsumi, T.; Eslam, M.; Kawaguchi, T.; Yamamura, S.; Kawaguchi, A.; Nakano, D.; Koseki, M.; Yoshinaga, S.; Takahashi, H.; Anzai, K.; et al. MAFLD better predicts the progression of atherosclerotic cardiovascular risk than NAFLD: Generalized estimating equation approach. Hepatol. Res. 2021, 51, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.A.; Lymp, J.F.; Sauver, J.S.; Sanderson, S.O.; Lindor, K.D.; Feldstein, A.; Angulo, P. The natural history of nonalcoholic fatty liver disease: A population-based cohort study. Gastroenterology 2005, 129, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.T.; Gidener, T.; Mara, K.C.; Larson, J.J.; Therneau, T.M.; Allen, A.M. Natural history of nonalcoholic fatty liver disease with normal body mass index: A population-based study. Clin. Gastroenterol. Hepatol. 2022, 20, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Golabi, P.; Paik, J.; Fukui, N.; Locklear, C.T.; de Avilla, L.; Younossi, Z.M. Patients with lean nonalcoholic fatty liver disease are metabolically abnormal and have a higher risk for mortality. Clin. Diabetes Publ. Am. Diabetes Assoc. 2019, 37, 65–72. [Google Scholar] [CrossRef] [PubMed]
- McDonagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Bohm, M.; Burri, H.; Butler, J.; Celutkiene, J.; Chinocel, O.; et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2021, 42, 3599–3726. [Google Scholar] [CrossRef] [PubMed]
- Van Riet, E.E.; Hoes, A.W.; Wagenaar, K.P.; Limburg, A.; Landman, M.A.; Rutten, F.H. Epidemiology of heart failure: The prevalence of heart failure and ventricular dysfunction in older adults over time. A systematic review. Eur. J. Heart Fail. 2016, 18, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.; Masson, S.; Anstee, Q.M. The bidirectional impacts of alcohol consumption and the metabolic syndrome: Cofactors for progressive fatty liver disease. J. Hepatol. 2018, 68, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Oben, J.A.; Roskams, T.; Yang, S.; Lin, H.; Sinelli, N.; Li, Z.; Torbenson, M.; Thomas, S.A.; Diehl, A.M. Norepinephrine induces hepatic fibrogenesis in leptin deficient ob/ob mice. Biochem. Biophys. Res. Commun. 2003, 308, 284–292. [Google Scholar] [CrossRef]
- Oben, J.A.; Diehl, A.M. Sympathetic nervous system regulation of liver repair. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 280, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fujimiya, T. Abrupt termination of an ethanol regimen provokes ventricular arrhythmia and enhances susceptibility to the arrhythmogenic effects of epinephrine in rats. Alcohol Clin. Exp. Res. 2010, 34 (Suppl. S1), S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Takase, I.; Hakucho, A.; Okamura, N.; Fujimiya, T. Carvedilol attenuates the progression of alcohol fatty liver disease in rats. Alcohol Clin. Exp. Res. 2012, 36, 1587–1599. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Alcohol consumption combined with dietary low-carbohydrate/high-protein intake increased the left ventricular systolic dysfunction risk and lethal ventricular arrhythmia susceptibility in apolipoprotein E/low-density lipoprotein receptor double-knockout mice. Alcohol 2020, 89, 63–74. [Google Scholar] [PubMed]
- Ejima, C.; Kuroda, H.; Ishizaki, S. A novel diet-induced murine model of steatohepatitis with fibrosis for screening and evaluation of drug candidates for nonalcoholic steatohepatitis. Physiol. Rep. 2016, 4, e13016. [Google Scholar] [CrossRef]
- Cheitlin, M.D.; Armstrong, W.F.; Aurigemma, G.P.; Beller, G.A.; Bierman, F.Z.; Davis, J.L.; Douglas, P.S.; Faxon, D.P.; Gillam, L.D.; Kimball, T.R. ACC/AHA/ASE 2003 guideline update for the clinical application of echocardiography: Summary article. A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines (ACC/AHA/ASE Committee to Update the 1997 Guidelines for the Clinical Application of Echocardiography). J. Am. Soc. Echocardiogr. 2003, 16, 1091–1110. [Google Scholar] [PubMed]
- Furuta, Y.; Liu, J.; Himemiya-Hakucho, A.; Yoshimura, K.; Fujimiya, T. Alcohol Consumption in Combination with an Atherogenic Diet Increased Indices of Atherosclerosis in Apolipoprotein E/Low-Density Lipoprotein Receptor Double-Knockout Mice. Alcohol Clin. Exp. Res. 2019, 43, 227–242. [Google Scholar] [CrossRef]
- Gadd, V.L.; Skoien, R.; Powell, E.E.; Fagan, K.J.; Winterford, C.; Horsfall, L.; Irvine, K.; Clouston, A.D. The portal inflammatory infiltrate and ductular reaction in human nonalcoholic fatty liver disease. Hepatology 2014, 59, 1393–4105. [Google Scholar] [CrossRef]
- Erlinger, S. Do patients with nonalcoholic fatty liver disease die from their heart? Clin. Res. Hepatol. Gastroenterol. 2011, 35, 163–165. [Google Scholar] [CrossRef]
- Sheka, A.C.; Adeyi, O.; Thompson, J.; Hameed, B.; Crawford, P.A.; Ikramuddin, S. Nonalcoholic Steatohepatitis: A Review. JAMA 2020, 323, 1175–1183. [Google Scholar] [CrossRef]
- Liang, W.; Menke, A.L.; Driessen, A.; Koek, G.H.; Lindeman, J.H.; Stoop, R.; Havekes, L.M.; Kleemann, R.; van den Hoek, A.M. Establishment of a general NAFLD scoring system for rodent models and comparison to human liver pathology. PLoS ONE 2014, 9, e115922. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Oba, Y.; Seiko Yamano, S. Chronic ethanol consumption plus dietary atherogenic diet intake created metabolic steatohepatitis with advanced liver fibrosis in apolipoprotein E/low-density lipoprotein receptor double-knockout mice. Alcohol Clin. Exp. Res. 2022, 46, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Rice, B.A.; Naimi, T.S.; Long, M.T. Nonheavy alcohol use associates with liver fibrosis and nonalcoholic steatohepatitis in the Framingham Heart Study. Clin. Gastroenterol. Hepatol. 2022, 8, S1542–S3565. [Google Scholar] [CrossRef]
- Long, M.T.; Massaro, J.M.; Hoffmann, U.; Benjamin, E.J.; Naimi, T.S. Alcohol use is associated with hepatic steatosis among persons with presumed nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2020, 18, 1831–1841. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Recent advances in understanding liver fibrosis: Bridging basic science and individualized treatment concepts. F1000Res 2018, 7, F1000 Faculty Rev-921. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.A. End-points for drug treatment in NASH. Hepatol. Int. 2019, 13, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Janig, W. Sympathetic nervous system and inflammation: A conceptual view. Auton. Neurosci. 2014, 182, 4–14. [Google Scholar] [CrossRef]
- Cervi, A.L.; Lukewich, M.K.; Lomax, A.E. Neural regulation of gastrointestinal inflammation: Role of the sympathetic nervous system. Auton. Neurosci. 2014, 182, 83–88. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Chen, H.; Zhou, H.; Liu, Z.; Liu, Z.; Liu, Z.; Zhou, Y.; Zhou, X.; Yu, L.; et al. Sympathetic nervous system mediates cardiac remodeling after myocardial infarction in a circadian disruption model. Front. Cardiovasc. Med. 2021, 8, 668387. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiraishi, S.; Liu, J.; Saito, Y.; Oba, Y.; Nishihara, Y.; Yoshimura, S. A New Non-Obese Steatohepatitis Mouse Model with Cardiac Dysfunction Induced by Addition of Ethanol to a High-Fat/High-Cholesterol Diet. Biology 2024, 13, 91. https://doi.org/10.3390/biology13020091
Shiraishi S, Liu J, Saito Y, Oba Y, Nishihara Y, Yoshimura S. A New Non-Obese Steatohepatitis Mouse Model with Cardiac Dysfunction Induced by Addition of Ethanol to a High-Fat/High-Cholesterol Diet. Biology. 2024; 13(2):91. https://doi.org/10.3390/biology13020091
Chicago/Turabian StyleShiraishi, Seiji, Jinyao Liu, Yuki Saito, Yumiko Oba, Yuiko Nishihara, and Satomichi Yoshimura. 2024. "A New Non-Obese Steatohepatitis Mouse Model with Cardiac Dysfunction Induced by Addition of Ethanol to a High-Fat/High-Cholesterol Diet" Biology 13, no. 2: 91. https://doi.org/10.3390/biology13020091