
Microstructural Adaptation for Prey Manipulation in the Millipede Assassin Bugs (Hemiptera: Reduviidae: Ectrichodiinae)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. Sample Preparation for SEM
2.3. Feeding Behavior Observing
3. Results
3.1. Type and Distribution of Antennal Sensilla
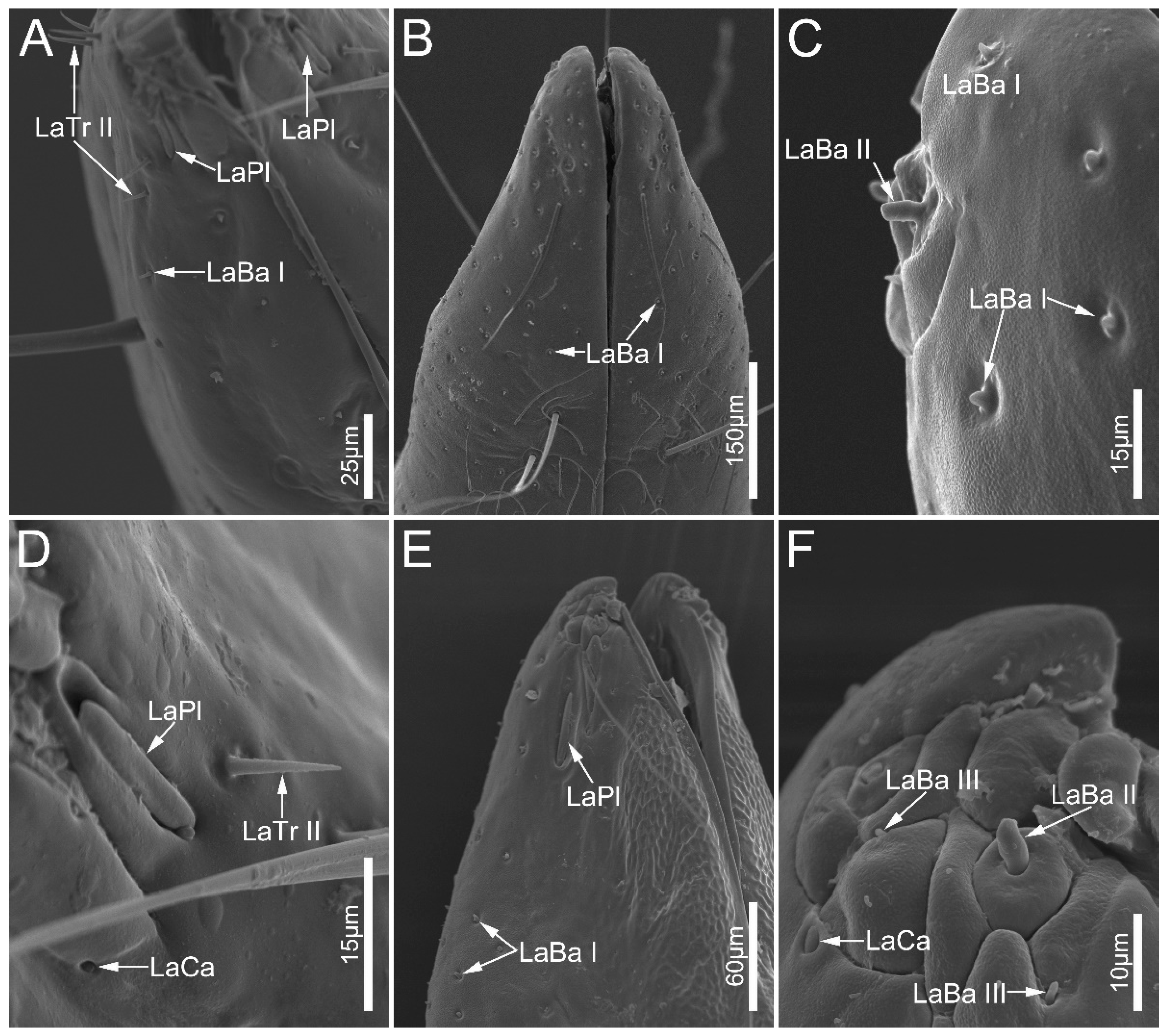
3.2. Type and Distribution of Labial Sensilla
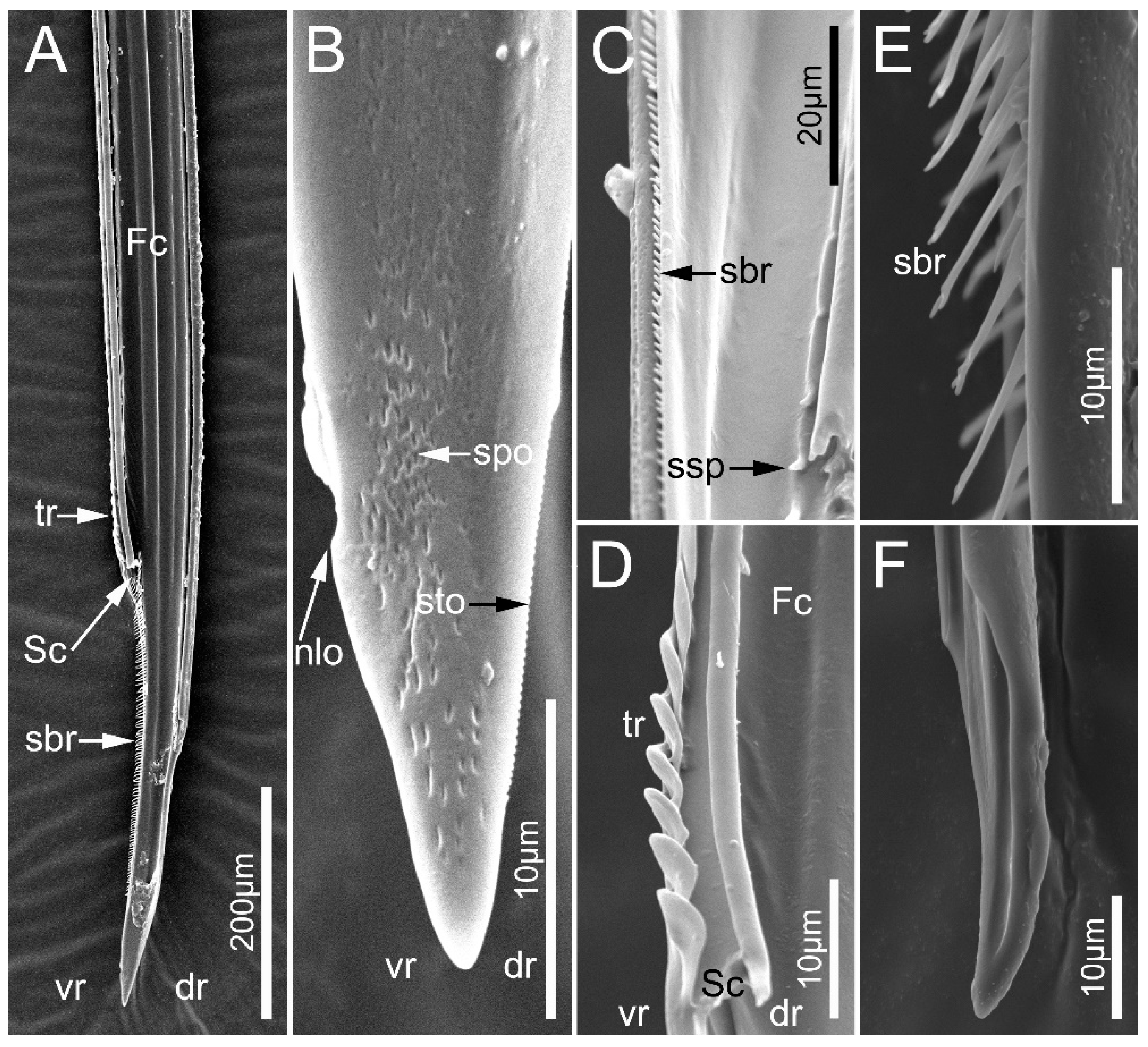
3.3. Fine Structure of Mandibles and Maxillae
3.4. Small Papillae on Legs
3.5. Predatory Behavior of Ectrichodiinae Species
4. Discussion
4.1. Morphological Adaptations of the Mouthparts for Millipede Feeding
4.2. Morphological Adaptation of the Legs for Millipede Feeding
4.3. Sensillar System on Antennae and Labium
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shear, W.A. The chemical defenses of millipedes (diplopoda): Biochemistry, physiology and ecology. Biochem. Syst. Ecol. 2015, 61, 78–117. [Google Scholar] [CrossRef]
- Weary, B.P.; Will, K.W.; Sime, K. The millipede-predation behavior of Promecognathus and exceptional cyanide tolerance in Promecognathus and Metrius (Coleoptera: Carabidae). Ann. Entomol. Soc. Am. 2020, 113, 473–480. [Google Scholar] [CrossRef]
- Tiemann, D. Observations on the natural history of the western banded glowworm, Zarhipis integripennis (Le Conte) (Coleoptera: Phengodidae). Proc. Calif. Acad. Sci. 1967, 35, 235–264. [Google Scholar]
- Eisner, T.; Eisner, M.; Attygalle, A.B.; Deyrup, M.; Meinwald, J. Rendering the inedible edible: Circumvention of a millipede’s chemical defense by a predaceous beetle larva (Phengodidae). Proc. Natl. Acad. Sci. USA 1998, 95, 1108–1113. [Google Scholar] [CrossRef]
- Forthman, M.; Weirauch, C. Toxic associations: A review of the predatory behaviors of millipede assassin bugs (Hemiptera: Reduviidae: Ectrichodiinae). Eur. J. Entomol. 2012, 109, 147–153. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Wang, W.; Brożek, J.; Dai, W. Unique fine morphology of mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) adapted to millipede feeding. Insects 2020, 11, 386. [Google Scholar] [CrossRef]
- Krell, F.-T. East African Dung Beetles (Scarabaeidae) Attracted by Defensive Secretions of Millipedes. J. East Afr. Nat. Hist. 2004, 93, 69–73. [Google Scholar] [CrossRef]
- Hash, J.M. Species of Megaselia Rondani (Diptera: Phoridae) attracted to defensive compounds of cyanogenic millipedes (Diplopoda: Polydesmida). Proc. Entomol. Soc. Wash. 2014, 116, 273–282. [Google Scholar] [CrossRef]
- Hash, J.M.; Millar, J.G.; Heraty, J.M.; Harwood, J.F.; Brown, B.V. Millipede Defensive Compounds Are a Double-Edged Sword: Natural History of the Millipede-Parasitic Genus Myriophora Brown (Diptera: Phoridae). J. Chem. Ecol. 2017, 43, 198–206. [Google Scholar] [CrossRef]
- Haridass, E.T.; Ananthakrishnan, T.N. Models for the predatory behavior of some reduviids from southern India (Insecta–Heteroptera–Reduviidae). Proc. Indian AS Anim. Sci. 1980, 89, 387–402. [Google Scholar] [CrossRef]
- Weirauch, C. Hairy attachment structures in Reduviidae (Cimicomorpha, Heteroptera), with observations on the fossula spongiosa in some other Cimicomorpha. Zool. Anz. 2007, 246, 155–175. [Google Scholar] [CrossRef]
- Weirauch, C. Cladistic analysis of Reduviidae (Heteroptera: Cimicomorpha) based on morphological characters. Syst. Entomol. 2008, 33, 229–274. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Weirauch, C. Sticky predators: A comparative study of sticky glands in harpactorine assassin bugs (Insecta: Hemiptera: Reduviidae). Acta. Zool 2013, 94, 1–10. [Google Scholar] [CrossRef]
- Zhang, J.; Gordon, E.R.; Forthman, M.; Hwang, W.S.; Walden, K.; Swanson, D.R.; Johnson, K.P.; Meier, R.; Weirauch, C. Evolution of the assassin’s arms: Insights from a phylogeny of combined transcriptomic and ribosomal DNA data (Heteroptera: Reduvioidea). Sci. Rep. 2016, 6, 22177. [Google Scholar] [CrossRef] [PubMed]
- Forthman, M.; Weirauch, C. Millipede assassins and allies (Heteroptera: Reduviidae: Ectrichodiinae, Tribelocephalinae): Total evidence phylogeny, revised classification and evolution of sexual dimorphism. Syst. Entomol. 2017, 42, 575–595. [Google Scholar] [CrossRef]
- Wang, J.Y. Taxonomic Study on the Subfamily Ectrichodiinae (Heteroptera: Reduviidae) from China and Regions around China. Ph.D. Thesis, China Agricultural University, Beijing, China, 2016. [Google Scholar]
- Global Biodiversity Information Facility. Opistoplatys majusculus Distant. 1904. Available online: https://www.gbif.org/species/9675201 (accessed on 18 September 2023).
- Global Biodiversity Information Facility. Tribelocephala walkeri China. 1940. Available online: https://www.gbif.org/zh/species/9812474 (accessed on 18 September 2023).
- Ambrose, D.P. Economic importance of Heteroptera. In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 695–712. [Google Scholar]
- Larsen, T.H.; Lopera, A.; Forsyth, A.; Genier, F. From coprophagy to predation: A dung beetle that kills millipedes. Biol. Lett. 2009, 5, 152–155. [Google Scholar] [CrossRef]
- Cobben, R.H. Evolutionary Trends in Heteroptera. Part II Mouthpart-Structures and Feeding Strategies; Mededelingen Land-Bouwhogeschool: Wageningen, The Netherlands, 1978; Volume 78, pp. 1–407. [Google Scholar]
- Schneider, D. Insect antennae. Annu. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Morphological disparity of the mouthparts in polyphagous species of Largidae (Heteroptera: Pentatomomorpha: Pyrrhocoroidea) reveals feeding specialization. Insects 2020, 11, 145. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Functional morphology and sexual dimorphism of antennae of the pear lace bug Stephanitis nashi (Hemiptera: Tingidae). Zool. Anz. 2020, 286, 11–19. [Google Scholar] [CrossRef]
- Li, X.; Tian, L.; Li, H.; Cai, W. Ultrastructural variations of antennae and labia are associated with feeding habit shifts in stink bugs (Heteroptera: Pentatomidae). Biology 2021, 10, 1161. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. Spirobolus bungii Brandt. 1833. Available online: https://www.gbif.org/species/1027616 (accessed on 18 September 2023).
- Global Biodiversity Information Facility. Orthomorphella pekuensis (Karsch, 1881). Available online: https://www.gbif.org/species/6127331 (accessed on 18 September 2023).
- Global Biodiversity Information Facility. Helicorthomorpha holstii (Pocock, 1895). Available online: https://www.gbif.org/species/1017372 (accessed on 18 September 2023).
- Global Biodiversity Information Facility. Oxidus gracilis (Koch, 1847). Available online: https://www.gbif.org/species/1017419 (accessed on 18 September 2023).
- Robert, P. Determinants of host-specificity in parasites of freshwater fishes. Int J. Parasitol 1992, 22, 753–758. [Google Scholar] [CrossRef]
- Pekar, S.; Coddington, J.A.; Blackledge, T.A. Evolution of stenophagy in spiders (Araneae): Evidence based on the comparative analysis of spider diets. Evolution 2012, 66, 776–806. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.S.; Weirauch, C. Evolutionary history of assassin bugs (insecta: Hemiptera: Reduviidae): Insights from divergence dating and ancestral state reconstruction. PLoS ONE 2012, 7, e45523. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.R.; Salm, K.; Nelson, X.J. Specialized prey selection behavior of two East African assassin bugs, Scipinnia repax and Nagusta sp. that prey on social jumping spiders. J. Insect Sci. 2010, 10, 82. [Google Scholar] [CrossRef]
- Wignall, A.E.; Taylor, P.W. Predatory behaviour of an araneophagic assassin bug. J. Ethol. 2010, 28, 437–445. [Google Scholar] [CrossRef]
- Hernandez, M.; Masonick, P.; Weirauch, C. Crowdsourced online images provide insights into predator-prey interactions of putative natural enemies. Food Webs 2019, 21, e00126. [Google Scholar] [CrossRef]
- Li, H.; Leavengood, J.M., Jr.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.; Zhou, X.; Cai, W. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. Biol. Sci. 2017, 284, 20171223. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.B.; Dai, W. Sensory armature and stylets of the mouthparts of Stephanitis nashi (Hemiptera: Cimicomorpha: Tingidae), their morphology and function. Micron 2020, 132, 102840. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. Fine structure of mouthparts and feeding performance of Pyrrhocoris sibiricus Kuschakevich with remarks on the specialization of sensilla and stylets for seed feeding. PLoS ONE 2017, 12, e0177209. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Comparative morphology of the mouthparts in three predatory stink bugs (Heteroptera: Asopinae) reveals feeding specialization of stylets and sensilla. Insects 2020, 11, 762. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. How does the intricate mouthpart apparatus coordinate for feeding in the hemimetabolous insect pest Erthesina fullo? Insects 2020, 11, 503. [Google Scholar] [CrossRef]
- Tull, T.; Henn, F.; Betz, O.; Eggs, B. Structure and function of the stylets of hematophagous Triatominae (Hemiptera: Reduviidae), with special reference to Dipetalogaster maxima. Arthropod. Struct. Dev. 2020, 58, 100952. [Google Scholar] [CrossRef] [PubMed]
- Bhagyasree, S.N.; Khader Khan, H. Morphological adaptations of forelegs associated with prey capture in assassin bugs (Reduviidae: Heteroptera). Mysore J. Agric. Sci. 2016, 50, 223–228. [Google Scholar]
- Dixon, J.W.; Rasmussen, R.D. Repeated predation of meadow fritillaries (Boloria bellona) by ambush bugs (Phymata spp.) (Note). J. Lepid. Soc. 2013, 67, 57–59. [Google Scholar] [CrossRef]
- Castro-Huertas, V.; Forero, D.; Grazia, J. Comparative morphology of the raptorial leg in thread-legged bugs of the tribe Metapterini Stål, 1859 (Hemiptera, Heteroptera, Reduviidae, Emesinae). Zoomorphology 2019, 138, 97–116. [Google Scholar] [CrossRef]
- Nowińska, A.F.-P.B.; Polhemus, D.A. The leg sensilla of insects from different habitats—Comparison of strictly aquatic and riparian bugs (Corixidae, Ochteridae, Gelastocoridae: Nepomorpha: Insecta: Heteroptera). Insects 2023, 14, 441. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Parveen, S.; Brożek, J.; Dey, D. Antennal sensilla of phytophagous and predatory pentatomids (Hemiptera: Pentatomidae): A comparative study of four genera. Zool. Anz. 2016, 261, 48–55. [Google Scholar] [CrossRef]
- Brézot, P.; Tauban, D.; Renou, M. Sense organs on the antennal flagellum of the green stink bug, Nezara viridula (L.) (Heteroptera: Pentatomidae) sensillum types and numerical growth during the post-embryonic development. Int. J. Insect Morphol. Embryol. 1997, 25, 427–441. [Google Scholar] [CrossRef]
- Sinitsina, E.E.; Chaika, S.Y. The Antenna sensory apparatus of terrestrial bugs (Heteroptera). Zool. Zhurnal 1998, 77, 1273–1284. [Google Scholar]
- Ventura, M.U.; Panizzi, A.R. Morphology of olfactory sensilla and its role in host plant recognition by Neomegalotomus parvus (Westwood) (Heteroptera: Alydidae). Braz. Arch. Biol. Technol. 2005, 48, 589–597. [Google Scholar] [CrossRef]
- Silva, C.C.; de Capdeville, G.; Moraes, M.C.; Falcao, R.; Solino, L.F.; Laumann, R.A.; Silva, J.P.; Borges, M. Morphology, distribution and abundance of antennal sensilla in three stink bug species (Hemiptera: Pentatomidae). Micron 2010, 41, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Weirauch, C. Pedicellar structures in Reduviidae (Heteroptera)—comments on cave organ and trichobothria. Eur. J. Entomol. 2003, 100, 571–580. [Google Scholar] [CrossRef]
- Taszakowski, A.; Gorczyca, J.; Herczek, A. Comparative study of the cephalic trichobothria in plant bugs (Hemiptera: Heteroptera: Miridae). Micron 2020, 137, 102918. [Google Scholar] [CrossRef] [PubMed]
- Taszakowski, A.; Masłowski, A.; Daane, K.M.; Brożek, J. Closer view of antennal sensory organs of two Leptoglossus species (Insecta, Hemiptera, Coreidae). Sci. Rep. 2023, 137, 102918. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.T.; Chen, X.; Ma, P.W.K. Labial tip sensilla of Blissus leucopterus leucopterus (Hemiptera: Blissidae): Ultrastructure and behavior. Insect Sci. 2008, 15, 271–275. [Google Scholar] [CrossRef]
- Parveen, S.; Ahmad, A.; Brożek, J.; Ramamurthy, V.V. Morphological diversity of the labial sensilla of phytophagous and predatory Pentatomidae (Hemiptera: Heteroptera), with reference to their possible functions. Zootaxa 2015, 4039, 359–372. [Google Scholar] [CrossRef]
- Brożek, J.; Chłond, D. Morphology, arrangement and classification of sensilla on the apical segment of labium in Peiratinae (Hemiptera: Heteroptera: Reduviidae). Zootaxa 2010, 2476, 39–52. [Google Scholar] [CrossRef]
- Taszakowski, A.; Maslowski, A.; Brożek, J. Labial sensory organs of two Leptoglossus species (Hemiptera: Coreidae): Their morphology and supposed function. Insects 2023, 14, 30. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Dai, W. Fine morphology of the mouthparts in Cheilocapsus nigrescens (Hemiptera: Heteroptera: Miridae) reflects adaptation for phytophagous habits. Insects 2019, 10, 43. [Google Scholar] [CrossRef]
- Taszakowski, A.; Nowińska, A.; Brożek, J. Morphological study of the labial sensilla in Nabidae (Hemiptera: Heteroptera: Cimicomorpha). Zoomorphology 2019, 138, 483–492. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Localization | Shape | Tip | Sockets |
|---|---|---|---|---|
| AnBa | flagellum | peg-shaped | blunt | inflexible |
| AnCh | pedicel, basiflagellomere, distiflagellomere | straight and stiff | sharp | flexible |
| AnTr I | pedicel, basiflagellomere, distiflagellomere | hair-shaped | tapered and sharp | inflexible |
| AnTr II | pedicel, basiflagellomere, distiflagellomere | hair-shaped | sharp | inflexible |
| AnTr III | all antennal segments | hair-shaped | sharp | inflexible |
| AnTb | pedicel | hair-shaped | sharp | set in a circular depression |
| Type | Localization | Shape | Sockets | Tip | Observed in Species |
|---|---|---|---|---|---|
| LaBa I | labial tip | peg-shaped | inflexible | blunt | all sampled species |
| LaBa II | labial tip | peg-shaped | inflexible | blunt | all sampled species |
| LaBa III | labial tip | peg-shaped | inflexible | blunt | Neozirta eidmanni |
| LaCa | ventral surface of labial tip | oval and dome-shaped | / | / | all sampled ectrichodiinae species |
| LaPl | lateral sides of the labial tip | dome-shaped | / | / | all sampled species |
| LaTr I | margin of labial apex | hair-shaped | flexible | sharp | all sampled species |
| LaTr II | labial apex | hair-shaped | inflexible | sharp | all sampled species |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zha, S.; Wang, Z.; Li, X.; Chen, Z.; Wang, J.; Li, H.; Cai, W.; Tian, L. Microstructural Adaptation for Prey Manipulation in the Millipede Assassin Bugs (Hemiptera: Reduviidae: Ectrichodiinae). Biology 2023, 12, 1299. https://doi.org/10.3390/biology12101299
Zha S, Wang Z, Li X, Chen Z, Wang J, Li H, Cai W, Tian L. Microstructural Adaptation for Prey Manipulation in the Millipede Assassin Bugs (Hemiptera: Reduviidae: Ectrichodiinae). Biology. 2023; 12(10):1299. https://doi.org/10.3390/biology12101299
Chicago/Turabian StyleZha, Shiyu, Zhiyao Wang, Xinyu Li, Zhaoyang Chen, Jianyun Wang, Hu Li, Wanzhi Cai, and Li Tian. 2023. "Microstructural Adaptation for Prey Manipulation in the Millipede Assassin Bugs (Hemiptera: Reduviidae: Ectrichodiinae)" Biology 12, no. 10: 1299. https://doi.org/10.3390/biology12101299