
First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea


Murmansk Marine Biological Institute of the Russian Academy of Sciences (MMBI RAS), 183038 Murmansk, Russia
*
Authors to whom correspondence should be addressed.
Diversity 2024, 16(1), 72; https://doi.org/10.3390/d16010072
Submission received: 13 December 2023
/
Revised: 19 January 2024
/
Accepted: 20 January 2024
/
Published: 22 January 2024
(This article belongs to the Special Issue Diversity, Phylogeny and Evolutionary History of Cnidaria)
Abstract
:Sarsia tubulosa (M. Sars, 1835), a colonial hydrozoan of the family Corynidae, is common in the Barents Sea. This study provides new evidence that the species acts as an epibiont of the red king crab Paralithodes camtschaticus, a non-indigenous but remarkably abundant decapod crustacean of immense commercial value in the coastal regions of the sea. This finding adds to the current collection of identified epibionts on the red king crab and highlights the prospect of ongoing symbiotic relationships between the crab and the regional fauna. We also provide photographic evidence of S. tubulosa colonies and a comparative morphological evaluation with another closely related corynid, Coryne hincksi Bonnevie, 1898, previously found on the red king crab. The main differences between these two species are the location of the gonophores (reproductive organs) on the hydranths and the final stage of their development: in the form of free-floating medusae (S. tubulosa) or remaining sessile as sporosacs (C. hincksi). Other distinguishing features include variations in hydranth morphology, number and arrangement of tentacles, and structural development of gonophores.
Epibiosis constitutes a facultative symbiotic relationship involving two distinct organisms: the epibiont and the basibiont. Organisms classified as epibionts, during their sessile life phase, adhere to the exterior of a living substrate. Conversely, the basibionts serve as the living substrate, providing support for the epibionts [1]. This type of symbiotic relationship provides an excellent model for investigating patterns of biodiversity across different geographical landscapes at multiple scales. Investigating epibiotic relationships can enhance the understanding of the critical biological characteristics of basibionts [1,2].
Hydrozoans are sessile invertebrates that colonize solid substrates, both biotic and abiotic and natural and artificial [3,4]. The typical life cycle of hydrozoans in the order Anthoathecata, diagnosed by the absence of a hydrotheca (a goblet-shaped external exoskeleton that protects the body), includes a polyp stage, in which individuals remain attached to the substrate and a free-living medusa phase. Polyps, which are mainly colonial organisms, are connected at the base by root-like tubes called stolons [5,6,7]. Although hydroid colonies are sexually dimorphic, with gonophores (containing gametes) serving as their reproductive organs, they have the ability to reproduce asexually, which they successfully use. Members of the family Corynidae typically inhabit shallow waters, and some species are relatively abundant [5,6,7,8,9]. Small polyp colonies in corynids display reptant behavior, mostly propagating on other organisms or forming “meadows” on primary substrates, thereby assuming an ecological role by becoming microhabitats for other species [10,11]. Unfortunately, these smaller hydroids are frequently disregarded as mere epizoites, hence leading to an information deficit concerning their ecological characteristics and roles within benthic and pelagic communities [12,13]. Hydrozoans repeatedly attach to the carapace of decapod crustaceans in the Barents Sea [14,15].
The red king crab Paralithodes camtschaticus is a large and economically significant crustacean deliberately introduced into the Barents Sea in the 1960s [16]. By the mid-1990s, this species had established a new self-sustaining population, and by the early 2000s, this population had reached levels suitable for commercial exploitation. Norway and Russia formally commenced large-scale red king crab fisheries in 2002 and 2004, respectively [16].
Currently, this crab acts as a host and supports a variety of symbiotic and epibiotic species, including hydrozoans [17,18,19]. In particular, the list of species known to colonize red king crabs shows a yearly increase [17,18,20].
Sarsia tubulosa (M. Sars, 1835), a colonial hydrozoan within the Anthoathecata order and Corynidae family, is found mainly along the boreal Atlantic coasts of Europe and North America, including the Arctic regions and the North Pacific. The geographic range to the south of Europe reaches the English Channel; however, medusae occur in Mediterranean lagoons [5,6,7]. The distribution of this species in the higher latitudes of Russia is well documented [4,21], although no previous records indicate its status as an epibiont of the red king crab. However, according to our unpublished data, it has been recorded on a gastropod Neptunea sp. and on the great spider crab Hyas araneus in the Barents Sea. There is a substantial body of research dedicated to investigating the hydromedusae stage of S. tubulosa [22,23,24,25,26]. However, data regarding the morphological characteristics of the polypoid stage of this species [5,6,27,28] lack accompanying high-quality photographs of live specimens. In addition, it is difficult to properly identify infertile material [6].
Our study aims to document the first record of S. tubulosa on red king crabs from the coastal Barents Sea. Additionally, it provides quality photographs of the living colonies, their comprehensive description, and a morphological comparison to the closely related Coryne hincksi Bonnevie, 1898, another member of the Corynidae family previously found on red king crabs. Although S. tubulosa is a common species in the Barents Sea, it has not been previously documented as an epibiont of the red king crab. In contrast, C. hincksi, a less well-known species that is not included in Russian guides of hydrozoan fauna [4,21], has regularly been registered in the epibiotic community of this host [17,19]. Therefore, this comparison aims to address the gap in morphological features and differentiation between these two epibiont hydrozoans.
A total of 70 red king crabs were collected by divers during a coastal expedition in July 2022 in Zelenetskaya Bay, on the base of Murmansk Marine Biological Institute in Dalnie Zelentsy (Figure 1). Our designated study area is a semi-enclosed bay on the eastern coast of the Kola Peninsula [29].
Once captured, the red king crabs were transported to the lab, where a thorough biometric examination was performed, including measurements and weight estimations. Each specimen was also examined for sex, shell condition, and epibiont presence, in accordance with our previous research methods [20,30]. Crab shell-age categories were visually determined using standard post-molt indicators, including the degree of abrasion on the ventral carapace, legs, and coxopodites, as well as coloration and spine wear [31].
Our sample size comprised 35 males (size range of 68.8–184.0 mm CL, mean of 157.8 ± 3.3 mm) and 35 females (size range of 116.8–146.0 mm, mean of 131.7 ± 1.3 mm). The majority of males fell into the 140–150 mm CL size class (29%), while the most abundant female cohort measured 130–140 mm CL (40%).
Colonies of S. tubulosa were identified on three large mature male crabs (Table 1).
The prevalence of infestation was 4.3% (95% confidence interval, ranging from 0.9 to 12.0%). Each crab infested with S. tubulosa colonies had either an old or very old shell, indicating that they had molted at least one year ago or more before being captured by our divers. Considering the large size of the collected colonies and their maturity status, it can be assumed that the S. tubulosa colonies have been present for a period exceeding several months. This is further supported by the absence of this epibiont on crabs with new shells that had molted 3–4 months prior to the study.
After collection, live hydrozoan colonies from Crab 2 and Crab 3 (hereafter referred to as Colony 2 and Colony 3, respectively) were individually placed in 80 mL plastic containers filled with chilled seawater (temperature of 5 °C, salinity of 34 psu). Subsequently, their morphological characteristics were examined [5,6,7], and they were photographed under an MBS-10 stereomicroscope equipped with a camera.
The colonies are pinkish in color, with a height of 1.6 cm (colony 2, Figure 2a) and 2.7 cm (colony 3, Figure 2b).
They have long, slender, and sparsely branched hydrocaulus (Figure 2c), with the largest stem diameter observed in the distal part ranging between 0.16 and 0.20 mm. The stems are covered by perisarc with either smooth or undulated walls, with annulations observed primarily in the proximal, narrower portions of the stems. Hydranths of S. tubulosa can reach a height of 1.3 mm and a diameter of 0.25–0.35 mm when expanded. Their shapes vary depending on the degree of contraction. There are 13–16 capitate tentacles (ranging from 0.23 to 0.54 mm in length and 0.07 to 0.09 mm in diameter) scattered over the upper 1/2–2/3 part of hydranth, including an oral whorl consisting of 4–5 oral tentacles. Distal spherical thickening of tentacles bearing a concentrated reserve of stinging capsules (nematocysts) is 0.09–0.15 mm in diameter. The filiform tentacles are absent.
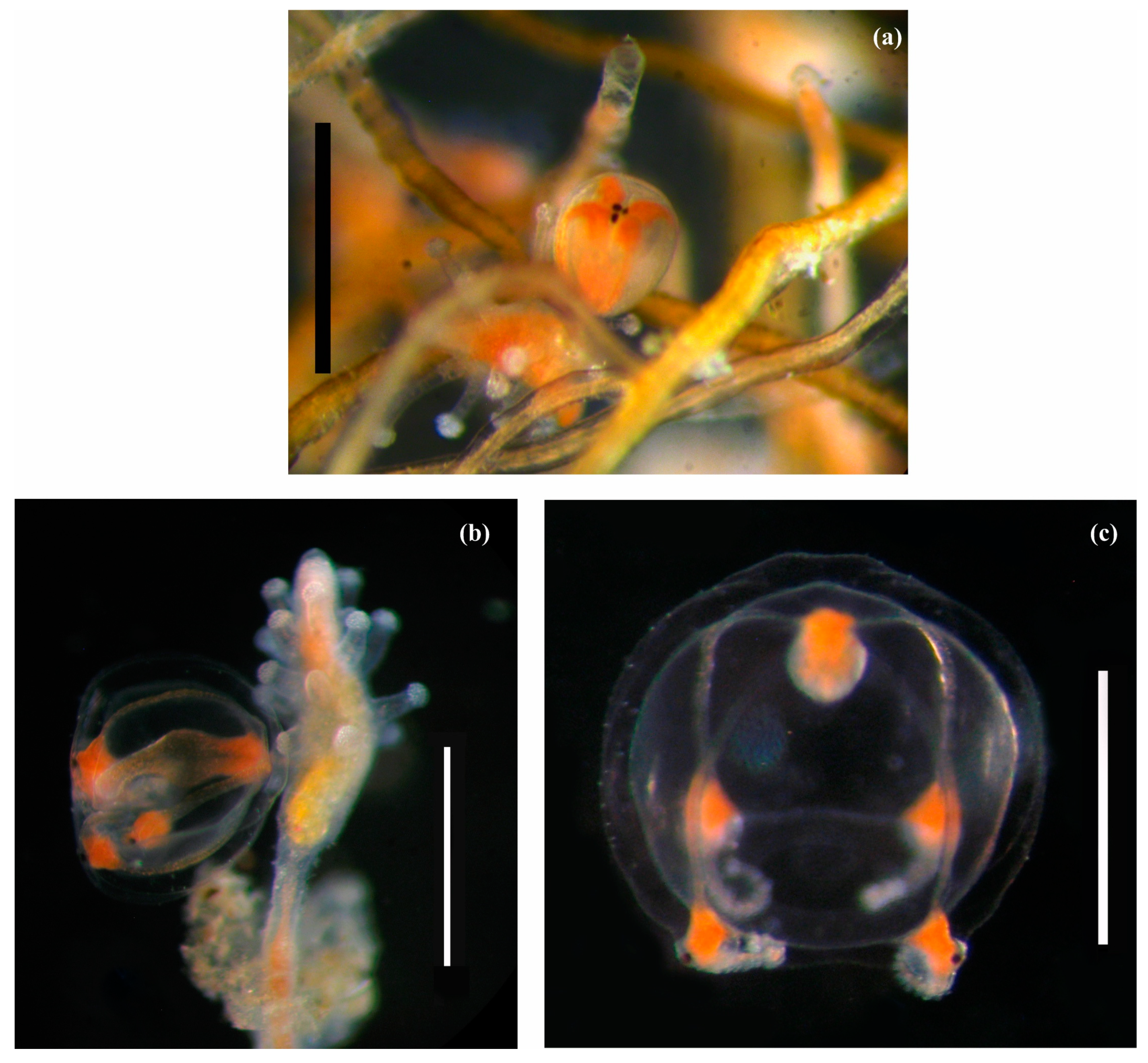
The gonophores develop into free-living medusae. Developing gonophores (medusoids) (typically 1–2 or possibly more per hydranth, being at different stages of maturation) are positioned on slender stalks (the stalk length does not exceed twice its thickness, 0.11 × 0.07 mm) below the lower tentacles or between them. Medusoids are spherical–cubic in shape, with four slender radial canals and four ruby–diamond-shaped tentacle bulbs. Each tentacle bulb hosts visible black eyes (Figure 3a). Fully developed medusoids (with a maximum diameter of up to 1.1 mm) have elongated tentacles concealed within the medusoid cavity (Figure 3b).
Before exiting the maternal colony, the fully developed medusoid displays pulsating activity and spreads its tentacles. The newly released medusa is spherical in shape (up to 1.5–1.65 mm in diameter) and has a short manubrium and a short, triangular apical chamber (Figure 2c).
At first glance, colonies of S. tubulosa appear very similar to Coryne hincksi, but the reproductive features of the two species differ significantly. The gonophores of S. tubulosa develop into free-living medusae, while those of C. hincksi are reduced to sessile sporosacs [19]. Furthermore, there are significant morphological differences between these two species. For example, the hydranths of C. hincksi are larger and have multiple tentacles scattered over the entire hydranth surface, appearing short and thick [19]. In contrast, the S. tubulosa hydranths are much smaller, with a smaller number of tentacles located in the upper half (or 2/3) of the polyp and appearing longer and thinner relative to the body of the polyp (Figure 4a).
The gonophores of S. tubulosa are located below the lowest tentacles or between them (at the same level) (Figure 4a), whereas those of C. hincksi are located between the tentacles in the lower half of the hydranth and can be at different levels (Figure 4b). In addition, developing medusoids of S. tubulosa have clearly visible black eyes (Figure 3a,b), whereas medusoids of C. hincksi lack eyes but have a wide, clearly visible ring canal at the distal end [19].
Colonies of S. tubulosa are present on solid substrates in the study area, regardless of their origin. In contrast, C. hincksi predominantly exhibits an epibiotic lifestyle, colonizing other invertebrates, particularly the bodies of spider crabs and lithodid crabs [19]. This association offers several advantages for corynid hydrozoans, such as protection against predators and increased mobility. Further research is necessary to determine if decapod hosts play a role in the life cycles of hydrozoans and other epibionts.
Author Contributions
Conceptualization, N.N.P. and A.G.D.; data curation, N.N.P., A.G.D., and V.G.D.; validation, N.N.P. and A.G.D.; methodology, N.N.P., V.G.D., and A.G.D.; software, A.G.D. and V.G.D.; visualization, N.N.P. and V.G.D.; project administration, V.G.D.; writing—original draft, N.N.P. and A.G.D.; writing—review and editing, N.N.P., A.G.D. and V.G.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Ministry of Science and Higher Education of the Russian Federation.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data are available on request from the corresponding author.
Acknowledgments
We are grateful to the academic editor and two anonymous reviewers for their valuable comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fernandez-Leborans, G. Epibiosis in Crustacea: An overview. Crustaceana 2010, 83, 549–640. [Google Scholar] [CrossRef]
- Williams, J.D.; McDermott, J.J. Hermit crab biocoenoses: A worldwide review of the diversity and natural history of hermit crab associates. J. Exp. Mar. Biol. Ecol. 2004, 305, 1–128. [Google Scholar] [CrossRef]
- Bouillon, J.; Gravili, C.; Pagès, F.; Gili, J.-M.; Boero, F. An introduction to hydrozoa. In Mémoires du Muséum National d’Histoire Naturelle; MarBEF: Aberdeen, Scotland, 2006; Volume 194, pp. 1–598. [Google Scholar]
- Antsulevich, A.E. Hydrozoa (Hydroids and Hydromedusae of Russian Seas); St. Petersburg University: St. Petersburg, Russia, 2015. (In Russian) [Google Scholar]
- Schuchert, P. Hydroids of Greenland (Cnidaria, Hydrozoa). Meddelelser om Gremland. Bioscience; Danish Polar Center: Copenhagen, Denmark, 2001; Volume 53. [Google Scholar]
- Schuchert, P. Survey of the family Corynidae (Cnidaria, Hydrozoa). Rev. Suisse Zool. 2001, 108, 739–878. [Google Scholar] [CrossRef]
- Schuchert, P. North-West European Athecate Hydroids and Their Medusae: Keys and Notes for the Identification of the Species; Dorset Press: Dorchester, UK, 2012. [Google Scholar]
- Rees, W.J. Evolutionary trends in the classification of capitate hydroids and medusae. Bull. Brit. Mus. Nat. Hist. Zool. 1957, 4, 453–534. [Google Scholar]
- Bouillon, J. Essai de classification des Hydropolypes−Hydroméduses (Hydrozoa-Cnidaria). Indo-Malayan Zool. 1985, 1, 29–243. [Google Scholar]
- Rees, J.T. The polyp and immature medusa stages of Cladonema californicum, Hyman, 1947 (Anthomedusae: Cladonemidae) with biological notes and a discussion of the taxonomy of the genus Cladonema. J. Nat. Hist. 1979, 13, 295–302. [Google Scholar] [CrossRef]
- Gili, J.M.; Lopez-Gonzalez, P.J.; Bouillon, J. A new Antarctic association: The case of the hydroid Sarsia medelae (new sp.) associated with gorgonians. Polar Biol. 2006, 29, 624–631. [Google Scholar] [CrossRef]
- Boero, F.; Gravili, C.; Denitto, F.; Miglietta, M.P.; Bouillon, J. The rediscovery of Codonorchis octaedrus (Hydroidomedusae, Anthomedusae, Pandeidae), with an update of the Mediterranean hydroidomedusan biodiversity. Ital. J. Zool. 1997, 64, 359–365. [Google Scholar] [CrossRef]
- Mills, C.E.; Marques, A.C.; Migotto, A.E.; Calder, D.R.; Hand, C.; Rees, J.T.S.; Haddock, H.D.C.; Dunn, W.; Pugh, P.R. Hydrozoa: Polyps, Hydromedusae, and Siphonophora. In The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon; Carlton, J.T., Ed.; University of California Press: Berkeley, CA, USA, 2007; pp. 118–167. [Google Scholar]
- Matishov, G.G. (Ed.) Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; KSC RAS: Apatity, Russia, 2008. (In Russian) [Google Scholar]
- Kuznetsov, V.V. The Biology of Abundant and the Most Common Species of Crustaceans in the Barents and White Seas; Nauka: Moscow, USSR, 1964. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Stevens, B.G. (Ed.) King Crabs of the World: Biology and Fisheries Management; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Panteleeva, N.N.; Dvoretsky, A.G.; Dvoretsky, V.G. New Records of the hydrozoan Coryne hincksi Bonnevie, 1898 on red king crabs in the Barents Sea. Diversity 2023, 15, 100. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef] [PubMed]
- Naumov, D.V. Hydroids and Hydromedusae of Marine, Brackish and Freshwater Basins of the USSR. In Guide to the Fauna of the USSR; Academy of Sciences: Moscow–Leningrad, USSR, 1960. (In Russian) [Google Scholar]
- Arai, M.N. Behavior of the planktonic coelenterates, Sarsia tubulosa, Phialidium gregarium, and Pleurobrachia pileus in salinity discontinuity layers. J. Fish. Board Can. 1973, 30, 1105–1110. [Google Scholar] [CrossRef]
- Singla, C.L.; Weber, C. Fine structure of the ocellus of Sarsia tubulosa (Hydrozoa, Anthomedusae). Zoomorphology 1982, 100, 11–22. [Google Scholar] [CrossRef]
- Leonard, J.L. The effect of environmental factors on swimming activity in Sarsia tubulosa M. Sars (Hydrozoa). Mar. Freshw. Behav. Physiol. 1983, 9, 99–110. [Google Scholar] [CrossRef]
- Daan, R. Food intake and growth of Sarsia tubulosa (Sars, 1835), with quantitative estimates of predation on copepod populations. Neth. J. Sea Res. 1986, 20, 67–74. [Google Scholar] [CrossRef]
- Sahin, M.; Mohseni, K.; Colin, S.P. The numerical comparison of flow patterns and propulsive performances for the hydromedusae Sarsia tubulosa and Aequorea victoria. J. Exp. Biol. 2009, 212, 2656–2667. [Google Scholar] [CrossRef]
- Edwards, C. The hydroids and medusae Sarsia occulta sp.nov., Sarsia tubulosa and Sarsia loveni. J. Mar. Biol. Assoc. UK 1978, 58, 291–311. [Google Scholar] [CrossRef]
- Bouillon, J.; Medel, M.D.; Pagès, F.; Gili, J.M.; Boero, F.; Gravili, C. Fauna of the Mediterranean hydrozoa. Sci. Mar. 2004, 68 (Suppl. S2), 5–438. [Google Scholar] [CrossRef]
- Il’in, G.V.; Moiseev, D.V.; Shirokolobov, D.V.; Deryabin, A.A.; Pavlova, L.G. Long-term dynamics of hydrological conditions of the Zelenetskaya Bay, East Murman. Vestn. MSTU 2016, 19, 268–277. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution of amphipods Ischyrocerus on the red king crab, Paralithodes camtschaticus: Possible interactions with the host in the Barents Sea. Estuar. Coast. Shelf Sci. 2009, 82, 390–396. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs. Fairbanks; Alaska Sea Grant College Program; University of Alaska: Fairbanks, AL, USA, 2005. [Google Scholar]
Figure 1.
Location of the study area (red dot) in the Barents Sea.

Figure 2.
Sarsia tubulosa (alive) from the Barents Sea. (a) Overall view of Colony 2, scale bar 5 mm; (b) Overall view of Colony 3, scale bar 5 mm; (c) Colony 3, scale bar 5 mm. Photo by Panteleeva, N.N.
Figure 2.
Sarsia tubulosa (alive) from the Barents Sea. (a) Overall view of Colony 2, scale bar 5 mm; (b) Overall view of Colony 3, scale bar 5 mm; (c) Colony 3, scale bar 5 mm. Photo by Panteleeva, N.N.

Figure 3.
Sarsia tubulosa (alive) from the Barents Sea. (a) Developing medusoid of Colony 3, scale bar 1 mm; (b) More developed medusoid of Colony 2, scale bar 1 mm; (c) Newly released medusa of Colony 2, scale bar 1 mm. Photo by Panteleeva, N.N.
Figure 3.
Sarsia tubulosa (alive) from the Barents Sea. (a) Developing medusoid of Colony 3, scale bar 1 mm; (b) More developed medusoid of Colony 2, scale bar 1 mm; (c) Newly released medusa of Colony 2, scale bar 1 mm. Photo by Panteleeva, N.N.

Figure 4.
Comparison of Sarsia tubulosa (a) and Coryne hincksi (b) live polyps with developing gonophores, showing differences in the size and distribution of tentacles and location of gonophores on hydranths. Scale bars 1 mm. Photo by Panteleeva, N.N.
Figure 4.
Comparison of Sarsia tubulosa (a) and Coryne hincksi (b) live polyps with developing gonophores, showing differences in the size and distribution of tentacles and location of gonophores on hydranths. Scale bars 1 mm. Photo by Panteleeva, N.N.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Information regarding findings and localization of Sarsia tubulosa on male red king crabs in Zelenetskaya Bay, southern Barents Sea, July 2022. Biological data of crabs (size, weight, and shell condition) are provided.
Table 1.
Information regarding findings and localization of Sarsia tubulosa on male red king crabs in Zelenetskaya Bay, southern Barents Sea, July 2022. Biological data of crabs (size, weight, and shell condition) are provided.
| No. | Date | Latitude, N | Longitude, E | Depth | T, °C | CL, mm | Wet Weight, g | Shell Condition | Localization |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 6 July 2022 | 69°07′10″ | 36°05′42″ | 18 | 4.0 | 156.0 | 3262 | Very old | Limbs |
| 2 | 12 July 2022 | 69°07′07″ | 36°04′24″ | 31 | 5.0 | 175.0 | 4676 | Old | Carapace |
| 3 | 13 July 2022 | 69°07′30″ | 36°04′48″ | 16 | 6.0 | 156.3 | 2643 | Old | Abdomen |
Note: T—water temperature, CL—carapace length; shell conditions as described in Donaldson and Byesrdorfer [31].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Panteleeva, N.N.; Dvoretsky, A.G.; Dvoretsky, V.G. First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea. Diversity 2024, 16, 72. https://doi.org/10.3390/d16010072
AMA Style
Panteleeva NN, Dvoretsky AG, Dvoretsky VG. First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea. Diversity. 2024; 16(1):72. https://doi.org/10.3390/d16010072
Chicago/Turabian StylePanteleeva, Ninel N., Alexander G. Dvoretsky, and Vladimir G. Dvoretsky. 2024. "First Record of Sarsia tubulosa (M. Sars, 1835) (Cnidaria, Hydrozoa) on Red King Crabs in the Coastal Barents Sea" Diversity 16, no. 1: 72. https://doi.org/10.3390/d16010072
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.