
Prolonged Differentiation of Neuron-Astrocyte Co-Cultures Results in Emergence of Dopaminergic Neurons
 , and
, and
Abstract
:1. Introduction
2. Results
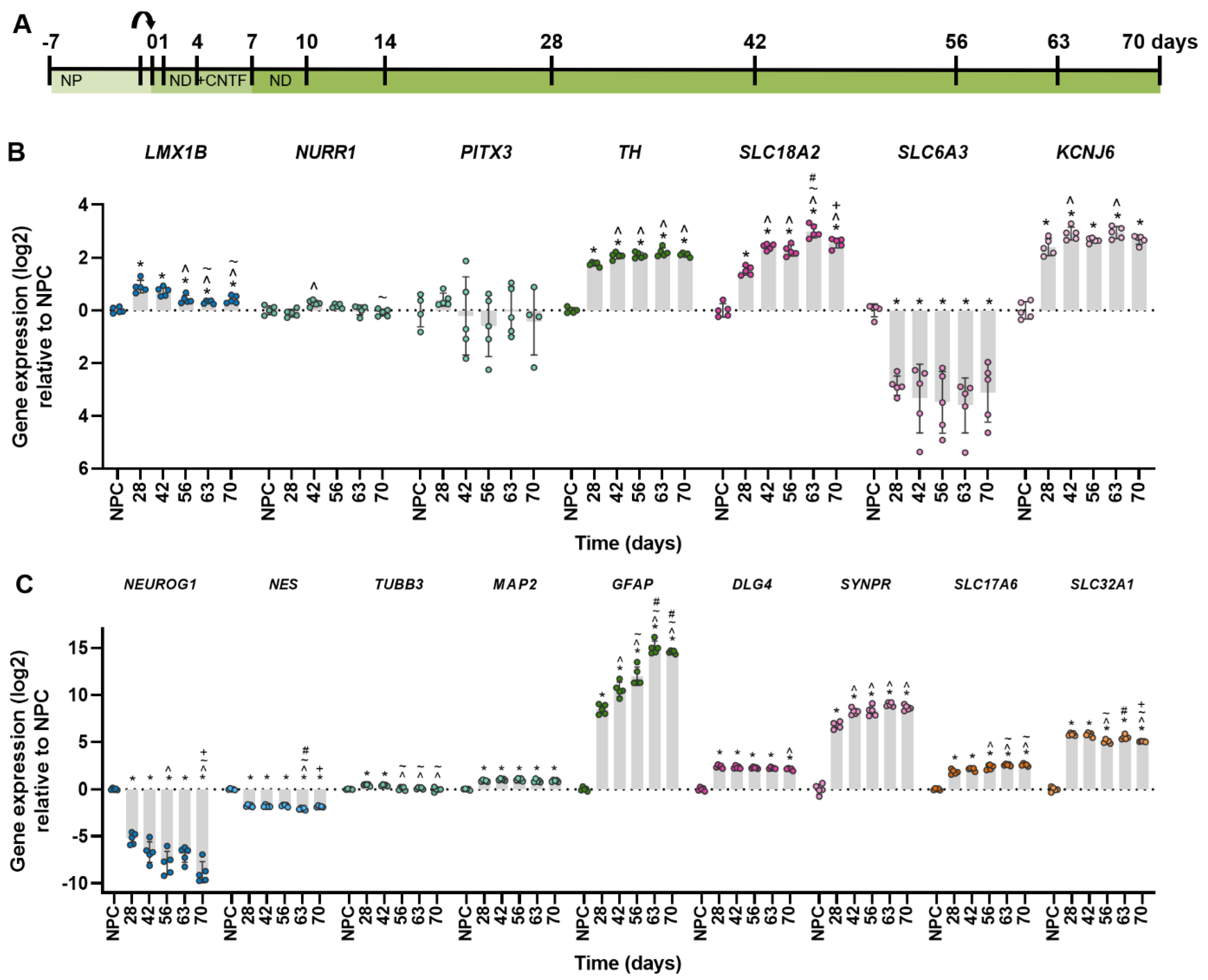
2.1. Robust Gene Expression of Dopaminergic Markers after Four Weeks of Differentiation
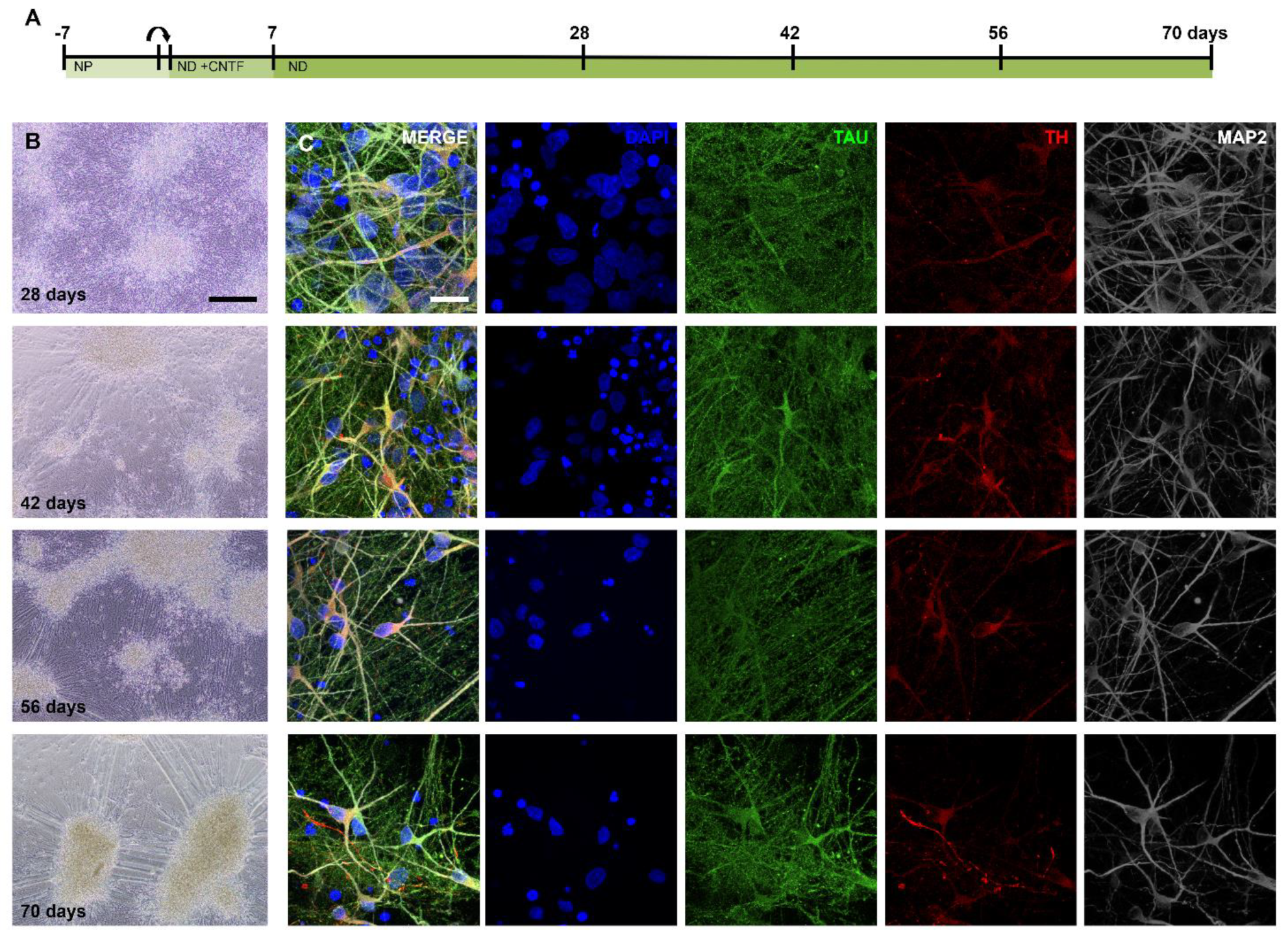
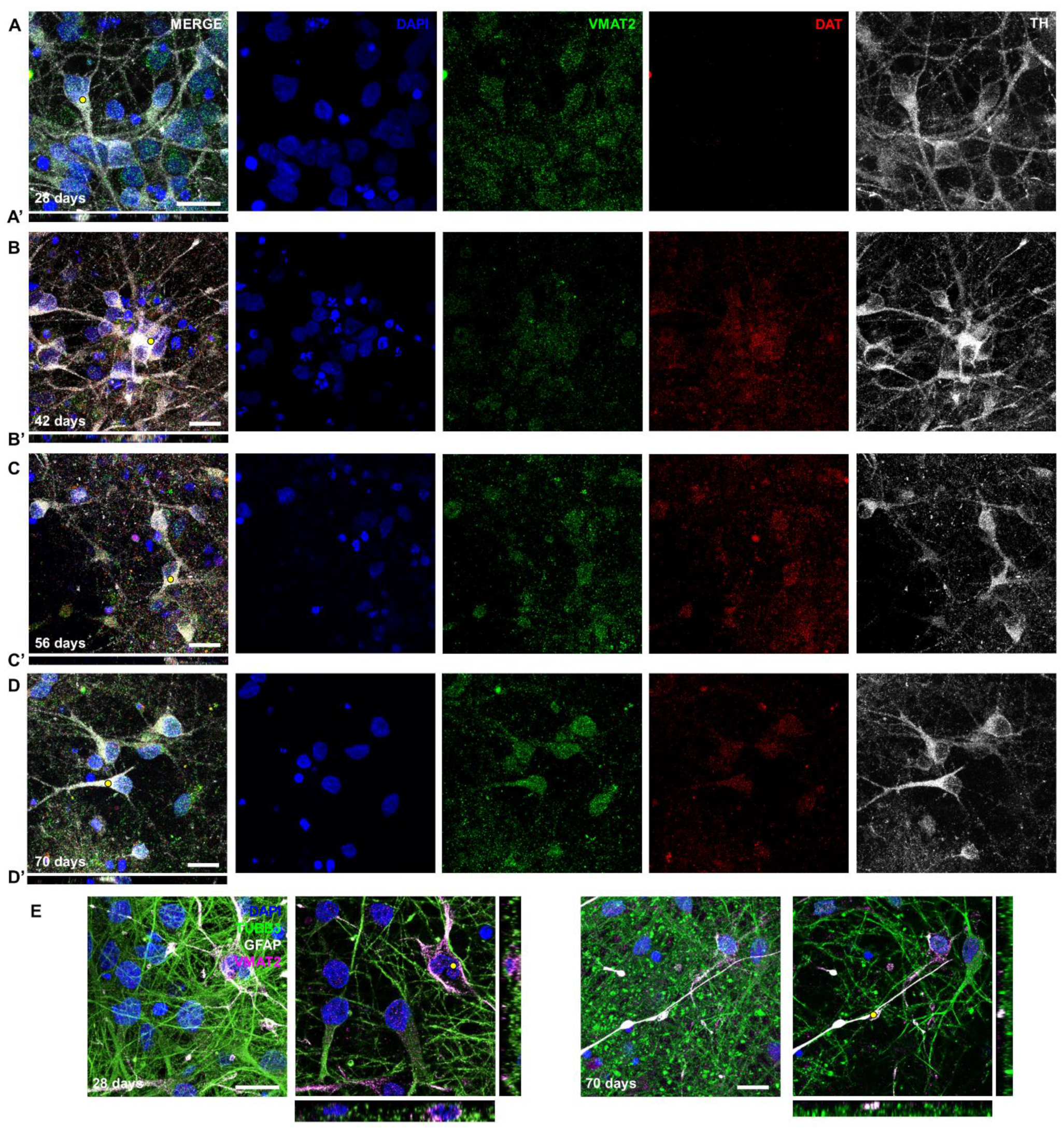
2.2. Generating Dopaminergic Neurons after Six Weeks of Differentiation
3. Discussion
4. Materials and Methods
4.1. Growth and Differentiation of a Neuron—Astrocyte Culture That Contains Dopaminergic Neurons from Embryonic Stem Cell-Derived Neural Progenitor Cells
4.2. Characterisation of Dopaminergic Neurons
4.3. RNA Isolation and qPCR
4.4. Immunocytochemistry
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, Y.; Xing, L.; Yang, T.; Chai, R.; Wang, J.; Bao, J.; Shen, W.; Ding, S.; Chen, G. The neurodevelopmental role of dopaminergic signaling in neurological disorders. Neurosci. Lett. 2021, 741, 135540. [Google Scholar] [CrossRef] [PubMed]
- Chaua, B.K.H.; Jarvisc, H.; Lawa, C.K.; Chongc, T.J. Dopamine and reward: A view from the prefrontal cortex. Behav. Pharmacol. 2018, 29, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Money, K.M.; Stanwood, G.D. Developmental origins of brain disorders: Roles for dopamine. Front. Cell. Neurosci. 2013, 7, 260. [Google Scholar] [CrossRef] [PubMed]
- Nikolaus, S.; Antke, C.; Müller, H.W. In vivo imaging of synaptic function in the central nervous system. I. Movement disorders and dementia. Behav. Brain Res. 2009, 204, 1–31. [Google Scholar] [CrossRef]
- Nikolaus, S.; Antke, C.; Müller, H.W. In vivo imaging of synaptic function in the central nervous system: II. Mental and affective disorders. Behav. Brain Res. 2009, 204, 32–66. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Burke, R.E.; O’Malley, K. Axon degeneration in Parkinson’s disease. Exp. Neurol. 2013, 246, 72–83. [Google Scholar] [CrossRef]
- Cheng, H.C.; Ulane, C.M.; Burke, R.E. Clinical progression in Parkinson disease and the neurobiology of axons. Ann. Neurol. 2010, 67, 715–725. [Google Scholar] [CrossRef]
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of Parkinson’s disease: Risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [Google Scholar] [CrossRef]
- Pang, S.Y.-Y.; Ho, P.W.-L.; Liu, H.-F.; Leung, C.-T.; Li, L.; Chang, E.E.S.; Ramsden, D.B.; Ho, S.-L. The interplay of aging, genetics and environmental factors in the pathogenesis of Parkinson’s disease. Transl. Neurodegener. 2019, 8, 23. [Google Scholar] [CrossRef]
- Hatcher, J.M.; Pennell, K.D.; Miller, G.W. Parkinson’s disease and pesticides: A toxicological perspective. Trends Pharmacol. Sci. 2008, 29, 322–329. [Google Scholar] [CrossRef]
- Moisan, F.; Spinosi, J.; Delabre, L.; Gourlet, V.; Mazurie, J.-L.; Bénatru, B.; Goldberg, M.; Weisskopf, M.G.; Imbernon, E.; Tzourio, C.; et al. Association of Parkinson’s Disease and Its Subtypes with Agricultural Pesticide Exposures in Men: A Case–Control Study in France. Environ. Health Perspect. 2015, 123, 1123–1129. [Google Scholar] [CrossRef]
- Narayan, S.; Liew, Z.; Bronstein, J.; Ritz, B. Occupational pesticide use and Parkinson’s disease in the Parkinson Environment Gene (PEG) study. Environ. Int. 2017, 107, 266–273. [Google Scholar] [CrossRef]
- Singh, C.; Ahmad, I.; Kumar, A. Pesticides and metals induced Parkinson’s disease: Involvement of free radicals and oxidative stress. Cell. Mol. Biol. 2007, 53, 19–28. [Google Scholar]
- van der Mark, M.; Vermeulen, R.; Nijssen, P.C.G.; Mulleners, W.M.; Sas, A.M.G.; van Laar, T.; Brouwer, M.; Huss, A.; Kromhout, H. Occupational exposure to pesticides and endotoxin and Parkinson disease in the Netherlands. Occup. Environ. Med. 2014, 71, 757LP–764. [Google Scholar] [CrossRef]
- Van Maele-Fabry, G.; Hoet, P.; Vilain, F.; Lison, D. Occupational exposure to pesticides and Parkinson’s disease: A systematic review and meta-analysis of cohort studies. Environ. Int. 2012, 46, 30–43. [Google Scholar] [CrossRef]
- Maitre, L.; Julvez, J.; López-Vicente, M.; Warembourg, C.; Tamayo-Uria, I.; Philippat, C.; Gützkow, K.B.; Guxens, M.; Andrusaityte, S.; Basagaña, X.; et al. Early-life environmental exposure determinants of child behavior in Europe: A longitudinal, population-based study. Environ. Int. 2021, 153, 106523. [Google Scholar] [CrossRef]
- Masjosthusmann, S.; Blum, J.; Bartmann, K.; Dolde, X.; Holzer, A.-K.; Stürzl, L.-C.; Keßel, E.H.; Förster, N.; Dönmez, A.; Klose, J.; et al. Establishment of an a priori protocol for the implementation and interpretation of an in-vitro testing battery for the assessment of developmental neurotoxicity. EFSA Support. Publ. 2020, 17, 1938E. [Google Scholar] [CrossRef]
- Vinken, M.; Benfenati, E.; Busquet, F.; Castell, J.; Clevert, D.-A.; de Kok, T.; Dirven, H.; Fritsche, E.; Geris, L.; Gozalbes, R.; et al. Safer chemicals using less animals: Kick-off of the European ONTOX project. Toxicology 2021, 458, 152846. [Google Scholar] [CrossRef]
- Hiller, B.M.; Marmion, D.J.; Thompson, C.A.; Elliot, N.A.; Federoff, H.; Brundin, P.; Mattis, V.B.; McMahon, C.W.; Kordower, J.H. Optimizing maturity and dose of iPSC-derived dopamine progenitor cell therapy for Parkinson’s disease. NPJ Regen. Med. 2022, 7, 24. [Google Scholar] [CrossRef]
- Kim, J.; Jeon, J.; Song, B.; Lee, N.; Ko, S.; Cha, Y.; Leblanc, P.; Seo, H. Spotting-based differentiation of functional dopaminergic progenitors from human pluripotent stem cells. Nat. Protoc. 2022, 17, 890–909. [Google Scholar] [CrossRef] [PubMed]
- Heusinkveld, H.J.; Westerink, R.H.S. Comparison of different in vitro cell models for the assessment of pesticide-induced dopaminergic neurotoxicity. Toxicol. Vitr. 2017, 45, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Meerman, J.J.; Wolterink, G.; Hessel, E.V.S.; de Jong, E.; Heusinkveld, H.J. Neurodegeneration in a Regulatory Context: The Need for Speed. Curr. Opin. Toxicol. 2022, 33, 100383. [Google Scholar] [CrossRef]
- Tebby, C.; Gao, W.; Delp, J.; Carta, G.; van der Stel, W.; Leist, M.; Jennings, P.; van de Water, B.; Bois, F.Y. A quantitative AOP of mitochondrial toxicity based on data from three cell lines. Toxicol. Vitr. 2022, 81, 105345. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, V.C.; van Oostrom, C.T.M.; Wackers, P.F.K.; Pennings, J.L.A.; Hodemaekers, H.M.; Piersma, A.H.; Hessel, E.V.S. Neuronal differentiation pathways and compound-induced developmental neurotoxicity in the human neural progenitor cell test (hNPT) revealed by RNA-seq. Chemosphere 2022, 304, 135298. [Google Scholar] [CrossRef]
- De Leeuw, V.C.; van Oostrom, C.T.M.; Westerink, R.H.S.; Piersma, A.H.; Heusinkveld, H.J.; Hessel, E.V.S. An efficient neuron-astrocyte differentiation protocol from human embryonic stem cell-derived neural progenitors to assess chemical-induced developmental neurotoxicity. Reprod. Toxicol. 2020, 98, 107–116. [Google Scholar] [CrossRef]
- Adil, M.M.; Rodrigues, G.M.C.; Kulkarni, R.U.; Rao, A.T.; Chernavsky, N.E.; Miller, E.W.; Schaffer, D.V. Efficient generation of hPSC-derived midbrain dopaminergic neurons in a fully defined, scalable, 3D biomaterial platform. Sci. Rep. 2017, 7, 40573. [Google Scholar] [CrossRef]
- Ásgrímsdóttir, E.S.; Arenas, E. Midbrain Dopaminergic Neuron Development at the Single Cell Level: In vivo and in Stem Cells. Front. Cell Dev. Biol. 2020, 8, 463. [Google Scholar] [CrossRef]
- OECD. Users’ Handbook Supplement to the Guidance Document for Developing and Assessing Adverse Outcome Pathways; OECD: Paris, France, 2018. [Google Scholar] [CrossRef]
- Van der Stel, W.; Carta, G.; Eakins, J.; Delp, J.; Suciu, I.; Forsby, A.; Cediel-Ulloa, A.; Attoff, K.; Troger, F.; Kamp, H.; et al. New Approach Methods (NAMs) Supporting Read-Across: Two Neurotoxicity AOP-based IATA Case Studies. ALTEX 2021, 38, 615–635. [Google Scholar] [CrossRef]
- Cao, Y. The uses of 3D human brain organoids for neurotoxicity evaluations: A review. Neurotoxicology 2022, 91, 84–93. [Google Scholar] [CrossRef]
- Pamies, D.; Barreras, P.; Block, K.; Makri, G.; Kumar, A.; Wiersma, D.; Smirnova, L.; Zhang, C.; Bressler, J.; Christian, K.M.; et al. A human brain microphysiological system derived from induced pluripotent stem cells to study neurological diseases and toxicity. ALTEX 2017, 34, 362–376. [Google Scholar] [CrossRef]
- Pistollato, F.; Mauro, P.; Clerbaux, L.A.; Leoni, G.; Ponti, J.; Bogni, A.; Brogna, C.; Cristoni, S.; Sanges, R.; Mendoza-de Gyves, E.; et al. Effects of spike protein and toxin-like peptides found in COVID-19 patients on human 3D neuronal/glial model undergoing differentiation: Possible implications for SARS-CoV-2 impact on brain development. Reprod. Toxicol. 2022, 111, 34–48. [Google Scholar] [CrossRef]
- Zagoura, D.; Canovas-Jorda, D.; Pistollato, F.; Bremer-Hoffmann, S.; Bal-Price, A. Evaluation of the rotenone-induced activation of the Nrf2 pathway in a neuronal model derived from human induced pluripotent stem cells. Neurochem. Int. 2017, 106, 62–73. [Google Scholar] [CrossRef]
- Gleichmann, M.; Mattson, M.P. Neuronal calcium homeostasis and dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Guzman, J.N.; Sanchez-Padilla, J.; Schumacker, P.T. The role of calcium and mitochondrial oxidant stress in the loss of substantia nigra pars compacta dopaminergic neurons in Parkinson’s disease. Neuroscience 2011, 198, 221–231. [Google Scholar] [CrossRef]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef]
- Krug, A.K.; Balmer, N.V.; Matt, F.; Schönenberger, F.; Merhof, D.; Leist, M. Evaluation of a human neurite growth assay as specific screen for developmental neurotoxicants. Arch. Toxicol. 2013, 87, 2215–2231. [Google Scholar] [CrossRef]
- Krug, A.K.; Gutbier, S.; Zhao, L.; Pöltl, D.; Kullmann, C.; Ivanova, V.; Förster, S.; Jagtap, S.; Meiser, J.; Leparc, G.; et al. Transcriptional and metabolic adaptation of human neurons to the mitochondrial toxicant MPP+. Cell Death Dis. 2014, 5, e1222. [Google Scholar] [CrossRef]
- Ransom, B.R.; Kunis, D.M.; Irwin, I.; Langston, J.W. Astrocytes convert the parkinsonism inducing neurotoxin, MPTP, to its active metabolite, MPP+. Neurosci. Lett. 1987, 75, 323–328. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, M.; Choi, I.; Ho, L.; Farrell, K.; Beaumont, K.G.; Sebra, R.; Crary, J.F.; Davis, D.A.; Sun, X.; et al. Single-cell Transcriptomic Atlas of the Human Substantia Nigra in Parkinson’s Disease. bioRxiv 2022, 2022, 485846. [Google Scholar] [CrossRef]
- Petrelli, F.; Dallérac, G.; Pucci, L.; Calì, C.; Zehnder, T.; Sultan, S.; Lecca, S.; Chicca, A.; Ivanov, A.; Asensio, C.S.; et al. Dysfunction of homeostatic control of dopamine by astrocytes in the developing prefrontal cortex leads to cognitive impairments. Mol. Psychiatry 2020, 25, 732–749. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, F.; Zehnder, T.; Pucci, L.; Cali, C.; Bondiolotti, B.M.; Perez, A.M.; Dallerac, G.; Déglon, N.; Giros, B.; Magara, F.; et al. Astrocytic VMAT2 in the developing prefrontal cortex is required for normal grooming behavior in mice. BioRvix 2021, 2021, 428434. [Google Scholar] [CrossRef]
- Agarwal, D.; Sandor, C.; Volpato, V.; Caffrey, T.M.; Monzón-Sandoval, J.; Bowden, R.; Alegre-Abarrategui, J.; Wade-Martins, R.; Webber, C. A single-cell atlas of the human substantia nigra reveals cell-specific pathways associated with neurological disorders. Nat. Commun. 2020, 11, 4183. [Google Scholar] [CrossRef] [PubMed]
- Gunhanlar, N.; Shpak, G.; van der Kroeg, M.; Gouty-Colomer, L.A.; Munshi, S.T.; Lendemeijer, B.; Ghazvini, M.; Dupont, C.; Hoogendijk, W.J.G.; Gribnau, J.; et al. A simplified protocol for differentiation of electrophysiologically mature neuronal networks from human induced pluripotent stem cells. Mol. Psychiatry 2018, 23, 1336–1344. [Google Scholar] [CrossRef]
- Applied Biosystems. User Bulletin #2 ABI PRISM 7700 Sequence Detection System; Applied Biosystems: Waltham, MA, USA, 2001. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Abbreviation | Marker for | Assay ID |
|---|---|---|---|
| Neurogenin 1 | NEUROG1 | Neural ectoderm | Hs01029249_s1 |
| Nestin | NES | Neural progenitor cell | Hs00707120_s1 |
| Tubulin, beta 3 class III | TUBB3 | Neuron | Hs00801390_s1 |
| Microtubule-associated protein 2 | MAP2 | Mature neuron | Hs00258900_m1 |
| Glial fibrillary acidic protein | GFAP | Early astrocyte | Hs00909233_m1 |
| Synaptoporin | SYNPR | Pre-synapse | Hs00376149_m1 |
| Discs Large MAGUK Scaffold Protein 4 | DLG4 | Post-synapse | Hs01555373_m1 |
| Vesicular glutamate transporter | SLC17A6 | Excitatory neuron | Hs00220439_m1 |
| Vesicular GABA transporter | SLC32A1 | Inhibitory neuron | Hs00369773_m1 |
| LIM homeobox transcription factor 1 beta | LMXB1 | Dopaminergic differentiation TF | Hs00158750_m1 |
| Paired Like homeodomain 3 | PITX3 | Dopaminergic differentiation TF | Hs00374504_m1 |
| Nuclear receptor subfamily 4 group A member 2 | NR4A2 | Dopaminergic differentiation TF | Hs00428691_m1 |
| Tyrosine Hydroxylase | TH | Catecholaminergic neuron | Hs00165941_m1 |
| Vesicular monoamine transporter 2 | VMAT2 | Monoamine transporter | Hs00996835_m1 |
| Dopamine transporter | SLC6A3 | Dopaminergic neuron | Hs00997374_m1 |
| Potassium voltage-gated channel subfamily J member 6 | KCNJ6 | Dopaminergic neuron | Hs00158423_m1 |
| Hypoxanthine phosphoribosyltransferase 1 | HPRT1 | Housekeeping gene | Hs02800695_m1 |
| RNA polymerase II subunit A | POLR2A | Housekeeping gene | Hs00172187_m1 |
| Glucuronidase beta | GUSB | Housekeeping gene | Hs00939627_m1 |
| Antibody | Abbreviation | Marker for | Product Number | Company | Dilution |
|---|---|---|---|---|---|
| Rabbit anti-β-Tubulin III | TUBB3 | Neuron | T2200 | Sigma-Aldrich | 1:1000 |
| Guinea-pig anti Tau | TAU | Neuon, axon | 314004 | Synaptic Systems | 1:1000 |
| Mouse anti-Microtubule-associated protein 2 | MAP2 | Neuron, dendrite | 801801 | Biolegend | 1:1000 |
| Rabbit anti-Tyrosine hydroxylase | TH | Catecholaminergic neuron | P40101-150 | Pel-Freez | 1:1000 |
| Rat anti-Glial fibrillary acidic protein | GFAP | Early astrocyte, radial glial cell | 13-0300 | Invitrogen | 1:800 |
| Mouse anti-Vesicular monoamine transporter 2 | VMAT2 | Monoamine transporter | MAB8327 | R&D Systems | 1:200 |
| Rat anti-Dopamine transporter | DAT | Dopaminergic neuron | AB-N17 | Advanced Targeting Systems | 1:350 |
| Goat anti-Rabbit Alexa 488 | A11034 | Invitrogen | 1:1000 | ||
| Goat anti-Guinea pig Alexa 488 | A11073 | Invitrogen | 1:1000 | ||
| Goat anti-Rabbit Alexa 555 | A21429 | Invitrogen | 1:1000 | ||
| Goat anti-Rat Alexa 555 | A21434 | Invitrogen | 1:500 | ||
| Goat anti-Mouse Alexa 647 | A21236 | Invitrogen | 1:500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Leeuw, V.C.; van Oostrom, C.T.M.; Zwart, E.P.; Heusinkveld, H.J.; Hessel, E.V.S. Prolonged Differentiation of Neuron-Astrocyte Co-Cultures Results in Emergence of Dopaminergic Neurons. Int. J. Mol. Sci. 2023, 24, 3608. https://doi.org/10.3390/ijms24043608
de Leeuw VC, van Oostrom CTM, Zwart EP, Heusinkveld HJ, Hessel EVS. Prolonged Differentiation of Neuron-Astrocyte Co-Cultures Results in Emergence of Dopaminergic Neurons. International Journal of Molecular Sciences. 2023; 24(4):3608. https://doi.org/10.3390/ijms24043608
Chicago/Turabian Stylede Leeuw, Victoria C., Conny T. M. van Oostrom, Edwin P. Zwart, Harm J. Heusinkveld, and Ellen V. S. Hessel. 2023. "Prolonged Differentiation of Neuron-Astrocyte Co-Cultures Results in Emergence of Dopaminergic Neurons" International Journal of Molecular Sciences 24, no. 4: 3608. https://doi.org/10.3390/ijms24043608